While it is not always obvious Loricariids all have teeth and they show a wide range of morphological disparity (Lujan & Armbruster, 2012). Morphological disparity refers to that range of different anatomy within a group. Unlike many fishes Loricariids are rarely gape limited, their prey (that includes algae) is not limited by the size of their mouth and this makes comparison with traditional fishes like carp or cichlids limited.
Figure 1: Pterygoplichthys joselimaianus jaw, a typical jaw for Loricariidae.
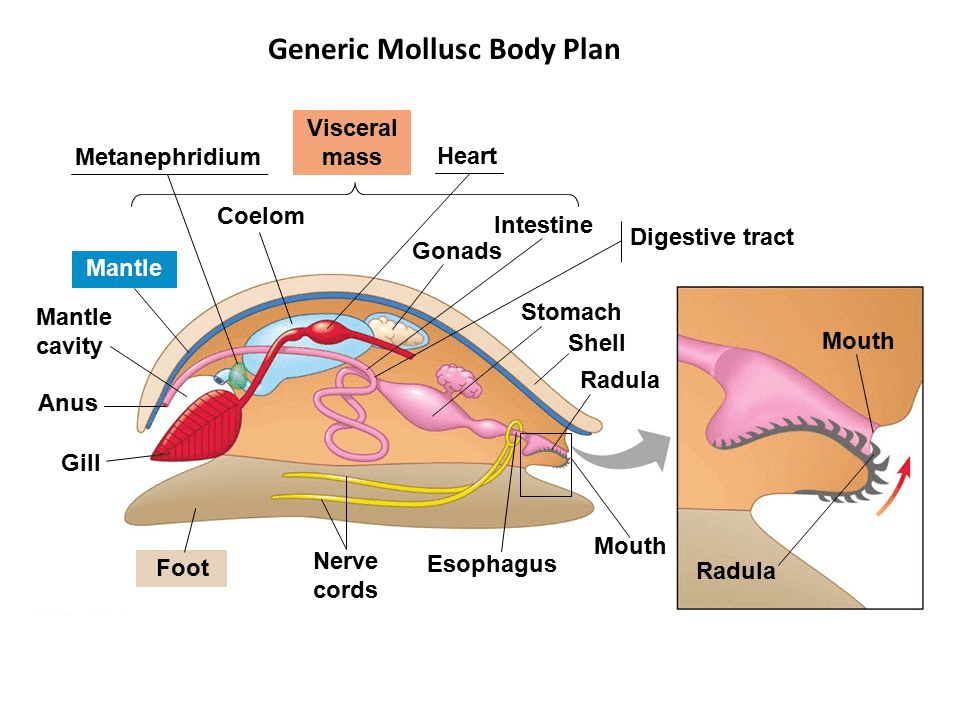
Loricariids feed largely by a rasping motion, this is extremely similar to how snails feed. Snails also have teeth on a ribbon like organ known as a radula (Fig 2).
Figure 2: Snail body plan including the mouth anatomy. Unknown source.
Both snails and Loricariids use their jaws, containing the teeth to basically scrape at a surface (rasp), it can be the food item itself but it could be rocks or wood to extract food.
Figure 3: Leporacanthicus joselimai
While I say the majority of Loricariid jaws are similar to snails not all are, there are carnivorous genera such as Pseudohemiodon or Scobinancistrus who differ in how they move and function. Carnivorous genera have elongate fewer teeth with often narrower oral jaws but can be much more robust (Fig 3), or in some those oral jaws are almost entirely just the jaws, reduced tooth cups. It’s easy to say these carnivores are using different morphology for the same solution to carnivory but maybe in a different place, one feeds amongst crevices (those with the elongate teeth and jaws) and others amongst the substrate (those with reduced jaws). We do have a slight exception with Spatuloricaria, an obvious substrate feeder but it seems to use the substrate a little differently and feed on different invertebrates.
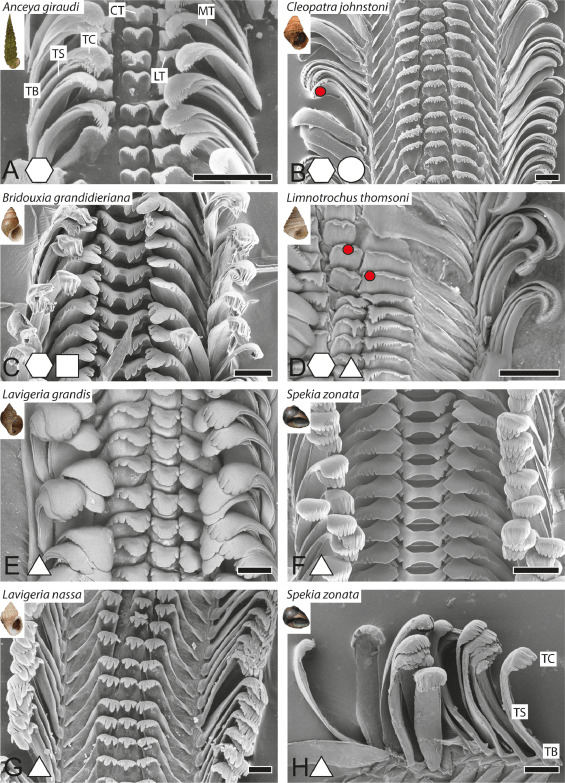
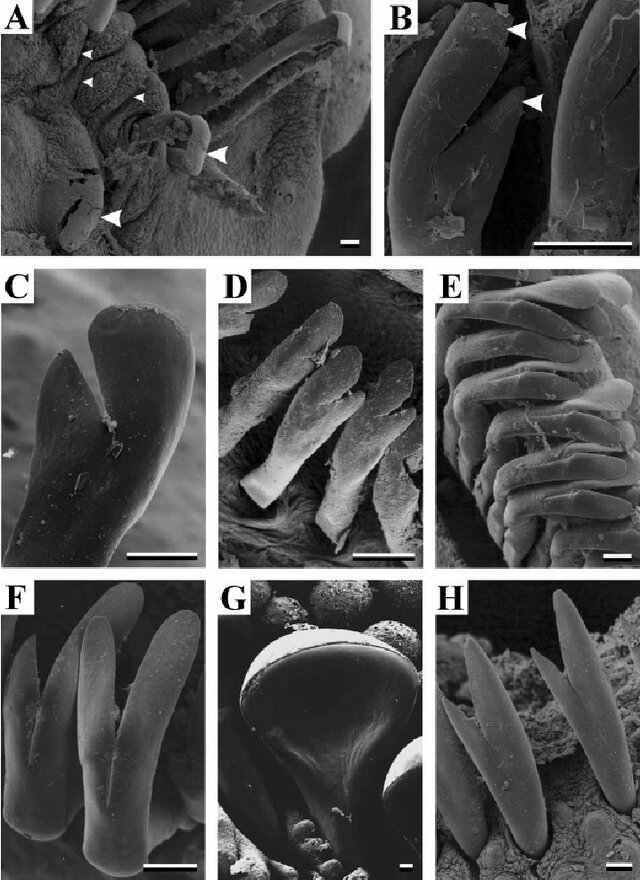
Figure 4: Gastropod radula diversity. Krings, W., Kovalev, A., & Gorb, S. N. (2021). Collective effect of damage prevention in taenioglossan radular teeth is related to the ecological niche in Paludomidae (Gastropoda: Cerithioidea). Acta Biomaterialia, 135, 458-472.
Snails, Gastropods have long been studied in terms of their radula diversity (Fig 4), I assume this is due to the fact when you’re dealing with preserved snails there are fewer tissues to identify the species. Additionally they make great models for understanding how anatomy relates to morphology, ecomorphology. Gastropods are everywhere and it’s easy to find those that scrape algae’s off rocks vs more carnivorous gastropods. To put it simply though, Gastropods feed by rasping and their teeth are uniquely shaped to what they are feeding on.
I think even just ignoring carnivory Loricariidae shows a wide diversity of tooth morphological disparity but there is little studies regarding that in relation to their ecology. Plenty of these studies focus on the development and morphology (Geerinckx et al., 2007). What there is is a fascinating study looking at another part of the fishes anatomy that could be similar, the unculi, small protrusions on the oral disc’s of the fishes. While the study focuses on how these structures allow for the fishes to inhabit certain habitats, could these also function in a similar fashion to radula?
Figure 5: The diversity of Loricariid teeth, Geerinckx, T., De Poorter, J., & Adriaens, D. (2007). Morphology and development of teeth and epidermal brushes in loricariid catfishes. Journal of morphology, 268(9), 805-814.
The diversity of Loricariid tooth morphological diversity is clear (Geerinckx et al., 2007) and we clearly see that Loricariids have a diversity of diets beyond herbivory and carnivory (Lujan et al., 2012), whatever they really mean to aquatic animals.
When looking outside of carnivory there is clear differences in morphology, none are so much clearer then those Loricariids that utilize wood. These genera display clearly spoon shaped teeth even if these genera (Panaqolus, Panaque, Hypostomus cochliodon group etc.) do not digest the wood and it is simply where they might find food. Compared with carnivores such as Leporacanthicus, these have more elongate teeth but it depends on where they are accessing their food. This difference is also reflected in gastropods whether they be snails or slugs have evolved teeth on their radula that reflect not just their diet but the methods they use to extract it. Elongate pointed teeth infers carnivory whereas further cusps leans towards herbivory. Perhaps carnivory requires less complexity to herbivory and I assume largely as carnivory relies on more then the teeth to extract food.
References:
Geerinckx, T., De Poorter, J., & Adriaens, D. (2007). Morphology and development of teeth and epidermal brushes in loricariid catfishes. Journal of morphology, 268(9), 805-814.
Krings, W., Konn-Vetterlein, D., Hausdorf, B., & Gorb, S. N. (2023). Holding in the stream: convergent evolution of suckermouth structures in Loricariidae (Siluriformes). Frontiers in Zoology, 20(1), 37.
Krings, W., Kovalev, A., & Gorb, S. N. (2021). Collective effect of damage prevention in taenioglossan radular teeth is related to the ecological niche in Paludomidae (Gastropoda: Cerithioidea). Acta Biomaterialia, 135, 458-472.
Lujan, N. K., & Armbruster, J. W. (2012). Morphological and functional diversity of the mandible in suckermouth armored catfishes (Siluriformes: Loricariidae). Journal of Morphology, 273(1), 24-39.
When you picture a freshwater ecosystem it really goes one of two ways, largely there is the group that look for a heavily planted water body with a diverse number of species or something that has a lot of leaf litter almost entirely complete for several layers.
Oase highline at Maidenhead Aquatics Ascot for Pseudancistrus asurini (Gold seam pleco), Baryancistrus xanthellus (Gold nugget pleco), Baryancistrus chrysolomus (Mango/magnum pleco) and rummynose tetra, Hemigrammus.
What if I told you this isn’t entirely true, a half truth or some what out of date?
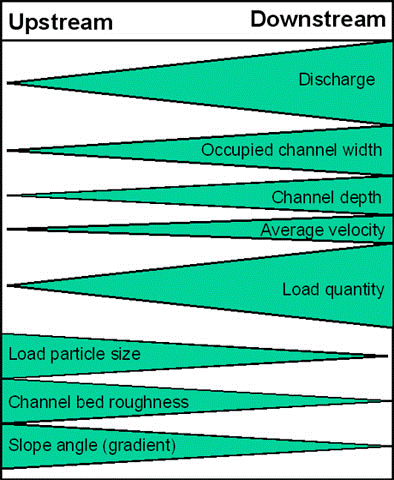
The Bradshaw model (Fig 1) explains this well but being largely based on temperate, North American and European ecosystems it doesn’t reflect tropical systems particularly as well.
The struggle for plant life.
When thinking about plants load particle size is important, plants in freshwater ecosystems largely need something to root onto and this isn’t found upstream where sediment is quickly pushed downstream. Downstream though this sediment load, load particle size is the opposite but because it’s remaining in the water column it becomes what is known as having a high turbidity (low visibility), this means less light can pass through the water and reduces photosynthesis. This is why so many aquarium plants are grown ‘terrestrially’ or aquaponically because they aren’t naturally aquatic. Aquatic plants have a struggle and there are solutions such as developing very strong roots and a reduced amount of structural support as if there is less sediment and a higher flow those stems are highly likely to break. In high turbidity waters that’s where we see the water lilies, Nymphaea spp., Nymphoides but also more floating plants who reach for the surface where they can receive light.
Some plants might have to deal with extreme drought in a dry season and then being drowned in the wet season. This is where we seem to see some really extremophile plants such as some Podostemaceae in the Rio Xingu likely of the genus Morera as reported by Peter Peterson in Amazonas (Fig 2).
Where is the leaf litter?
This is such a similar story for leaf litter as deposition, and particle size largely depends on flow rate but position in the river. Firstly you require that river to be surrounded by trees but also that the current is weak enough that the leaves can just fall to the bottom. When you look at rivers like the Rio Xingu, Rio Orinoco or towards the Rift Valley this is not shown in the major areas of the rivers or lakes where many of our fishes are caught. Hillstreams where other fishes are caught if leaves and fruits fall then the current might be so strong it’ll push it downstream to be deposited in certain areas of a meander known as deposition zones.
Aqua One 360, 60L for Pangio spp. and Apistogramma
This makes it just really complex to generalize but this is also why some areas have high tannin composition, Rio Negro and others have none such as the main channel of the Amazon and where they meet is known as the meeting of waters. The Rio Negro’s distinct black waters are likely due to it’s geology (Sioli, 1968), but likely those contributing tributaries cause a lot of the difference. Are these tributaries hillstream’s and likely to be less surrounded by plants at those higher elevations? Or maybe that geology largely contributes to the types of trees surrounding smaller source streams? There is a lot to unpack about why, but as hobbyists I’m not sure this why really matters? What matters is what these water bodies are and why what they are effects our fishes.
The reality of freshwater habitat diversity
It is a little bit of a pain but the image we have of freshwater habitats is curated by the idea perpetuated by brands and hobbyists. There is the idea that black water, therefore leaf litter is so common and when we look at the rivers and lakes around us, surely we must notice the difference?
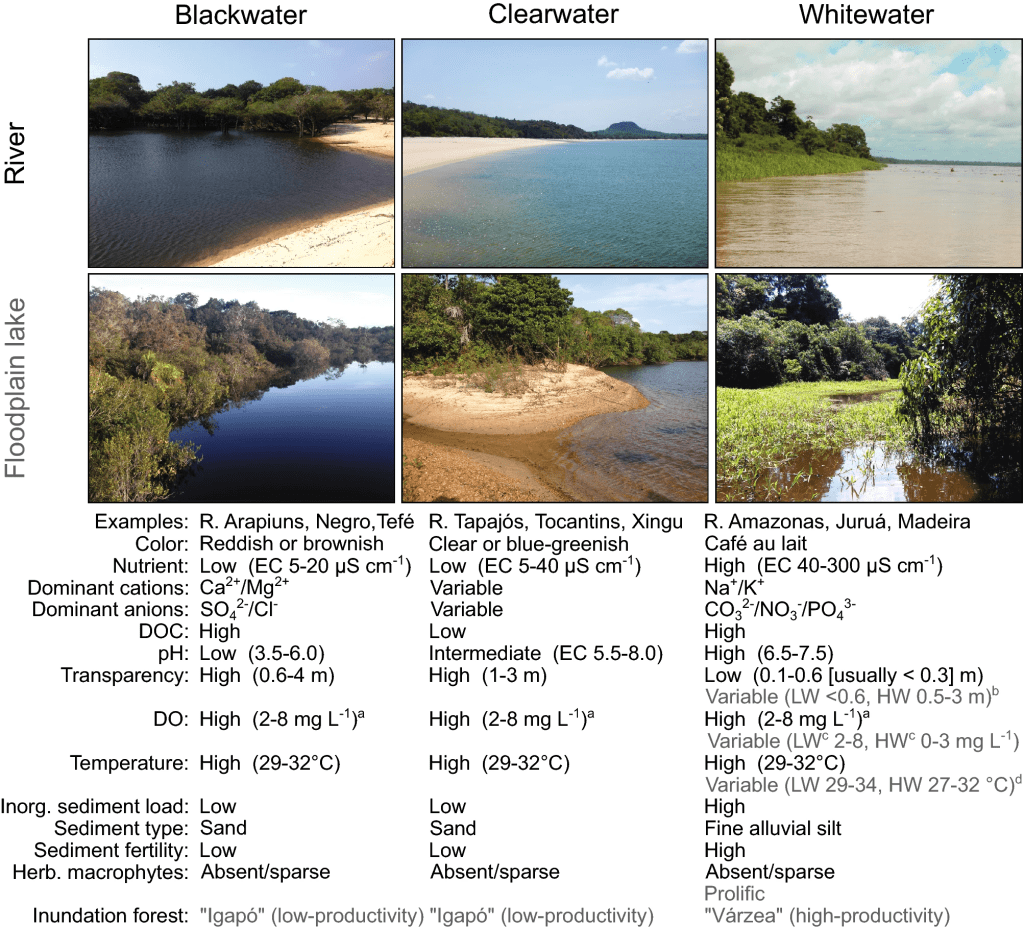
Rivers are generally classified into white water or black water and then subdivided further as explained so clearly by Bogotá-Gregory et al. (2020; fig. 3).
Figure 3: Classification of freshwater river types, can some what apply to lakes source: Bogotá-Gregory, J. D., Lima, F. C., Correa, S. B., Silva-Oliveira, C., Jenkins, D. G., Ribeiro, F. R., … & Crampton, W. G. (2020). Biogeochemical water type influences community composition, species richness, and biomass in megadiverse Amazonian fish assemblages. Scientific Reports, 10(1), 15349.
Do I need to say more? I wonder if the influence of these ideas of freshwater ecosystems comes from the passionate fishkeepers focused on those smaller niche fish such as wild Betta (not domestic Betta splendens), other anabantoids and what I some what try to affectionately say swimmy fishes. Previously these black water, leaf litter tanks would have been seen as dirty but they have effectively justified this ecosystem for the hobby. The problem has come where it’s shoved almost all freshwater species into being black water, particularly if from South America.
Lakes also show a wide amount of diversity between lakes and within that same lake there can be many different habitats.
The deception of anthropomorphic effects
As humans encroach on land and change it, the function changes too. This has been happening for thousands of years but regardless this effects the species that inhabit these waters. Rivers have been straightened and moved, stopped and redirected or even slowed and sped up. New lakes are created and others destroyed. Our waste thrown into rivers and lakes whether it be effluent or invasive species, warm water from coolant systems or mining waste. Freshwater is changing and fishes cannot always adapt fast enough. Your local river might not always be representative of nature, the plants are likely invasive and the sediment is likely due effluent or waste. The presence of a dam or weir means that the entire flow of the river has changed.
Figure 4: Habitat of Scobinancistrus raonii in the RIo Xingu as featured in: Chaves, M. S., Oliveira, R. R., Gonçalves, A. P., Sousa, L. M., & Py-Daniel, L. H. R. (2023). A new species of armored catfish of the genus Scobinancistrus (Loricariidae: Hypostominae) from the Xingu River basin, Brazil. Neotropical Ichthyology, 21(3), e230038.
This can be similar as to when we look at wild habitats, but much of the footage and images we see today seem to have been taken a while ago. Still the footage and photos of the Rio Xingu is showing that amount of sediment that might be connected to the building of the Belo Monte dam in 2011.
Source of nutrients
This is not where the debate is in the hobby but in the science it certainly is (Neres‐Lima et al., 2017; Hamilton et al., 1992). The long held belief in the hobby is that nutrients originates from leaf litter, fruit, branches etc. Therefore it’s important in the aquarium as some how all fishes even if there is no leaf litter in their habitat benefit from it.
This idea that allochthonous (leaf litter, fruit, branches, other botanicals etc.) nutrients is the major source of nutrients over autochthonous (algaes) can be true in some ecosystems but it doesn’t mean that nutrients is accessible (Thorp & Delong, 2002) and therefore few organisms obtain nutrients from it. This theory of nutrients deriving from allochthonous relies on the presence of leaf litter, botanicals being present in the water or further upstream and in shaded rivers it might contribute where no algae’s are found (Neres-lime et al., 2017). One answer answer is black water, often characterized by high botanicals/allochthonous influences is much lower in productivity and if a source of nutrients this should be another story (Bogotá-Gregory et al., 2020; Lewis et al., 1988). We also know as hobbyists algae’s can grow in very dimly lit aquariums though, nutrients is a whole other discussion.
It doesn’t seem an easy question to answer as while isotopes seem to be an answer and algae’s contribute most to nutrients in one study (Hamilton et al., 1992). As the cited papers suggest rivers flowing through slower flowing areas might contribute to nutrients opposed to it being directly in the rivers, but this means that the nutrients is in the source water not where the fishes are.
Why does algae do so well but plants don’t?
Algae’s are found in all of these habitats but to varying amounts, many do require light as photosynthetic organisms but some are known as heterotrophs so can gain energy by other means. It’s not just algae’s but other microbes that grow in these ecosystems, all of these unlike plants cling to surfaces and in a very small size meaning that there is much less drag. If they do grow too big then parts get broken off to throw downstream. What might be forgotten is bryophytes also do this but still photosynthetic. This leaves sponges as filter feeders.
What is algae?
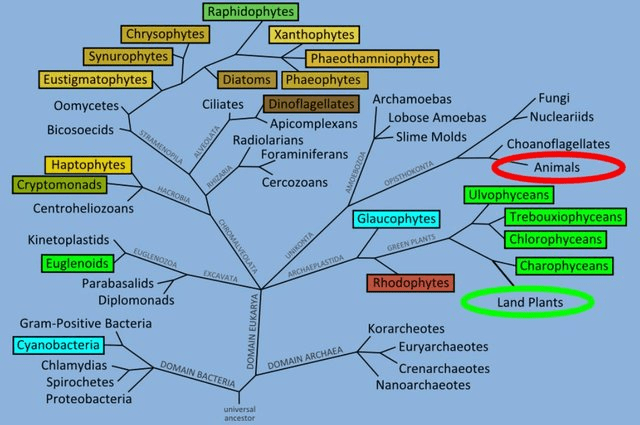
Figure 5: Phylogeny of algae source: SCHLARB-RIDLEY, B. 2011. Algal Research in the UK. A Report for Bbsrc. BBSRC. and Milledge, J. J. (2013). Energy balance and techno-economic assessment of algal biofuel production systems (Doctoral dissertation, University of Southampton).
Algae (Alga, singular) is basically a common name for a whole mixture of different organisms (Milledge, 2013). Some hobbyists will exclude Cyanobacteria as it is a bacteria yet it makes little sense given this clade is already polyphyletic (Fig 5), a real pick and mix. Another opinion is as these organisms are photosynthetic they must be similar to plants, plants particularly vegetables make good nutritional replacements. As you can see from the above it’s unlikely plants make up for algae’s nutritionally (Čengić-Džomba et al., 2022; Nagappan et al., 2021; Vucko et al., 2017).
This makes algae such a diverse grouping to describe as earlier stated some being heterotrophs do not just obtain their ‘energy’ from the sun and others might obtain it from feeding or even parasitism e.g. dinoflagellates (Levy et al., 2007; Sudhagar et al., 2022).
Algae is so important to an ecosystem and I will include other microbial life on that. This wide diversity of organisms feature in many fishes diets even making the majority of many Loricariid diets (Lujan et al., 2015). There is a link between algae’s and the fishes that feed on them influences the habitat’s and those fishes (Power, 1984).
Algae’s have to compete with different microbes and this is fascinating in terms of succession. Succession is the process of a habitat maturing over time with the change in species present. This happens in an aquarium over time but also how we maintain our aquariums seems to effect this. Generally you can expect as a grazing fish moves over an area it removes all the organisms it feeds best on, leaving an opening for other species of microbes to grow. These original fishes might not feed on that new microbes but other fishes will have and could this be where niche partitioning further occurs.
Long live algae! Leaf litter is overrated.
You don’t need a green aquarium to embrace them, realistically if you’re getting high amounts of algae it could infer high levels of a compound or element that encourages their growth as I keep stating. Above where I mention hypothetical niche partitioning regarding algae’s and other surface growing microbes. This will inevitably occur with bacterial films or any growths on leaf litter, if you’re seeing the films on botanicals in general for a while they probably aren’t being eaten by anything and when they eventually go it could be due to exhausting the source of nutrients. I particularly noted this when experimenting with different woods and palms, if these biofilms were being eaten there wouldn’t be so much biofilm’s growing. So far in Loricariids at least we know this niche partitioning is largely in where food items are located (Lujan et al., 2011) and while their diets aren’t the best studied we do have a realistic idea that they are not all feeding on the same algae’s or microbes (Lujan et al., 2012; Valencia & Zamudio, 2007; Delariva & Agostinho, 2001).
It is frequently stated that leaf litter has beneficial properties due to tannins but it is rarely mentioned that tannins are antinutrients and can cause physiological damage (Omnes et al., 2017; Li et al., 2020; Maulidiyah et al., 2019). Given some fishes inhabit these naturally they likely do have physiological mechanisms to manage tannins within the water, although given many fishes do have negative physiological responses as cited above not all do.
The frequent dislike of tea tree oils in the hobby is contrasted with the support for tannins having antimicrobial properties and they possibly do but in prevention and handling of disease in fishes, these effects are minimal (Imperatore et al., 2022). Perhaps the bias against tea tree oils is due to a dislike of brands as a review by Valladão et al. (2015) infers that use of known concentrations of compounds of plant origins (herbal treatments) can show very effective use. Leaf litter and tannins from the leaf litter have a flaw here, species identification and in using the plant as a whole it is difficult to say what compounds and concentrations are being added. We are living in an anti-pharmaceutical age and who can tell us more about how compounds interact with physiology then pharmacologists as the story is quite complex.
Leaf litter is not all bad
Leaf litter and botanicals can add great hiding spaces for smaller fishes, there is still a number of fishes who find it as a part of their natural habitat and this is worth replicating. The main downfall with leaf litter is siphoning to remove waste and therefore excess nutrients, other heavier botanicals would be more ideal if this is a big concern. But without evidence we shouldn’t jump to conclusions.
Pangio myersi with leaf litter, from Springs Aquatic Ltd, Botley, Hampshire.
Is there hope for algal growth?
No doubt algae does actually struggle to grow to much of an extent in a mature aquarium that isn’t constantly being scrubbed. For an aquarium which might benefit from ‘naturally’ growing algae’s such as for feeding fishes there is hope. We can’t use sunlight like outside but perhaps using UVB lighting could encourage growth. Most of our aquarium algae growth relies on nutrients but outside sunlight would perhaps contribute, something I’d certainly like to explore more.
Aquariums are anthropogenic
At the end of the day aquariums are anthropogenic and the argument about the importance of leaf litter vs algae really stall’s there. Algae’s are generally great indicators of nutrients, generally different algae’s hint to the age of an aquarium.
At the end maybe the promotion of leaf litter is easy to justify as natural then algae. There are few true biotopes and few ever show algae’s but the scale you’d need to grow them would be insane.
References:
Delariva, R. L., & Agostinho, A. A. (2001). Relationship between morphology and diets of six neotropical loricariids. Journal of Fish biology, 58(3), 832-847.
Bogotá-Gregory, J. D., Lima, F. C., Correa, S. B., Silva-Oliveira, C., Jenkins, D. G., Ribeiro, F. R., … & Crampton, W. G. (2020). Biogeochemical water type influences community composition, species richness, and biomass in megadiverse Amazonian fish assemblages. Scientific Reports, 10(1), 15349.
Čengić-Džomba, S., Džomba, E., & Hadžić, D. (2022). An Overview of Using Algae Meal in Feeding Freshwater Fish Species. In Scientific-Expert Conference of Agriculture and Food Industry (pp. 171-182). Cham: Springer Nature Switzerland.
Chaves, M. S., Oliveira, R. R., Gonçalves, A. P., Sousa, L. M., & Py-Daniel, L. H. R. (2023). A new species of armored catfish of the genus Scobinancistrus (Loricariidae: Hypostominae) from the Xingu River basin, Brazil. Neotropical Ichthyology, 21(3), e230038.
Farha, A. K., Yang, Q. Q., Kim, G., Li, H. B., Zhu, F., Liu, H. Y., … & Corke, H. (2020). Tannins as an alternative to antibiotics. Food Bioscience, 38, 100751.
Hamilton, S. K., Lewis, W. M., & Sippel, S. J. (1992). Energy sources for aquatic animals in the Orinoco River floodplain: evidence from stable isotopes. Oecologia, 89, 324-330.
Imperatore, R., Fronte, B., Scicchitano, D., Orso, G., Marchese, M., Mero, S., … & Paolucci, M. (2022). Dietary supplementation with a blend of hydrolyzable and condensed Tannins ameliorates diet-induced intestinal Inflammation in Zebrafish (Danio rerio). Animals, 13(1), 167.
Küchler, I. L., Miekeley, N., & Forsberg, B. R. (2000). A contribution to the chemical characterization of rivers in the Rio Negro Basin, Brazil. Journal of the Brazilian Chemical Society, 11, 286-292.
Levy, M. G., Litaker, R. W., Goldstein, R. J., Dykstra, M. J., Vandersea, M. W., & Noga, E. J. (2007). Piscinoodinium, a fish-ectoparasitic dinoflagellate, is a member of the class Dinophyceae, subclass Gymnodiniphycidae: convergent evolution with Amyloodinium. Journal of Parasitology, 93(5), 1006-1015.
Lewis Jr, W. M. (1988). Primary production in the Orinoco River. Ecology, 69(3), 679-692.
Li, M., Feng, L., Jiang, W. D., Wu, P., Liu, Y., Jiang, J., … & Zhou, X. Q. (2020). Condensed tannins decreased the growth performance and impaired intestinal immune function in on-growing grass carp (Ctenopharyngodon idella). British Journal of Nutrition, 123(7), 737-755.
Lujan, N. K., German, D. P., & Winemiller, K. O. (2011). Do wood‐grazing fishes partition their niche?: morphological and isotopic evidence for trophic segregation in Neotropical Loricariidae. Functional Ecology, 25(6), 1327-1338.
Lujan, N. K., Winemiller, K. O., & Armbruster, J. W. (2012). Trophic diversity in the evolution and community assembly of loricariid catfishes. BMC Evolutionary Biology, 12, 1-13.
Maulidiyah, V., Sulmartiwi, L., & Masithah, E. D. (2019). The effect of immersion time in tannin solution towards the adhesiveness and hatching degree of the eggs of common carp (Cyprinus carpio). AACL Bioflux.
Milledge, J. J. (2013). Energy balance and techno-economic assessment of algal biofuel production systems (Doctoral dissertation, University of Southampton).
Nagappan, S., Das, P., AbdulQuadir, M., Thaher, M., Khan, S., Mahata, C., … & Kumar, G. (2021). Potential of microalgae as a sustainable feed ingredient for aquaculture. Journal of Biotechnology, 341, 1-20.
Neres‐Lima, V., Machado‐Silva, F., Baptista, D. F., Oliveira, R. B., Andrade, P. M., Oliveira, A. F., … & Moulton, T. P. (2017). Allochthonous and autochthonous carbon flows in food webs of tropical forest streams. Freshwater Biology, 62(6), 1012-1023.
Omnes, M. H., Le Goasduff, J., Le Delliou, H., Le Bayon, N., Quazuguel, P., & Robin, J. H. (2017). Effects of dietary tannin on growth, feed utilization and digestibility, and carcass composition in juvenile European seabass (Dicentrarchus labrax L.). Aquaculture Reports, 6, 21-27.
Power, M. E. (1984). Habitat quality and the distribution of algae-grazing catfish in a Panamanian stream. The Journal of Animal Ecology, 357-374.
Sioli, H. (1968). Hydrochemlstry and Geology in the Brazilian Amazon Region. Volume 1, fascículo 3, 1968.
Sudhagar, A., Sundar Raj, N., Mohandas, S. P., Serin, S., Sibi, K. K., Sanil, N. K., & Raja Swaminathan, T. (2022). Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius). Pathogens, 11(11), 1350.
Thorp, J. H., & Delong, M. D. (2002). Dominance of autochthonous autotrophic carbon in food webs of heterotrophic rivers. Oikos, 96(3), 543-550.
Valencia, C. R., & Zamudio, H. (2007). Dieta y reproducción de Lasiancistrus caucanus (Pisces: Loricariidae) en la cuenca del río La Vieja, Alto Cauca, Colombia. Revista del Museo Argentino de Ciencias Naturales nueva serie, 9(2), 95-101.
Valladão, G. M. R., Gallani, S. U., & Pilarski, F. (2015). Phytotherapy as an alternative for treating fish disease. Journal of veterinary pharmacology and therapeutics, 38(5), 417-428.
Vucko, M. J., Cole, A. J., Moorhead, J. A., Pit, J., & de Nys, R. (2017). The freshwater macroalga Oedogonium intermedium can meet the nutritional requirements of the herbivorous fish Ancistrus cirrhosus. Algal research, 27, 21-31.
Cichlids, cichlidae are no doubt the classic model group for the study of evolution, they display a wide diversity of morphology in both the America’s and Africa (Arbour & López‐Fernández, 2024; Santos et al., 2023). There are 1727 species in the group cichlidae making this family currently the largest family of fishes. (Fricke et al., 2024). Cichlids both sides of the Atlantic are popular in the aquarium hobby, partially due to their diversity (no doubt colouration) but also their comparatively heightened aggression compared to other clades. No doubt aggression is where many people empathize with with these fishes.
Symphysodon tarzoo, the green discus but also includes some browns and blues.
What do cichlids specialize in?
Not unexpected, many species lack a complete understanding of their diet to say what they are actually feeding on. This is a vast topic given the number of species, there are algivores (algae specialists) such as reported in Tropheus (Wagner et al., 2009) to piscivory in peacock bass, Cichla (Aguiar‐Santos et al., 2018). No doubt given I said that Tropheus are reported algivores means I can’t find any solid evidence that they are, this is certainly a topic that needs the right wording and ambiguity included where it might be unsure.
Morphology
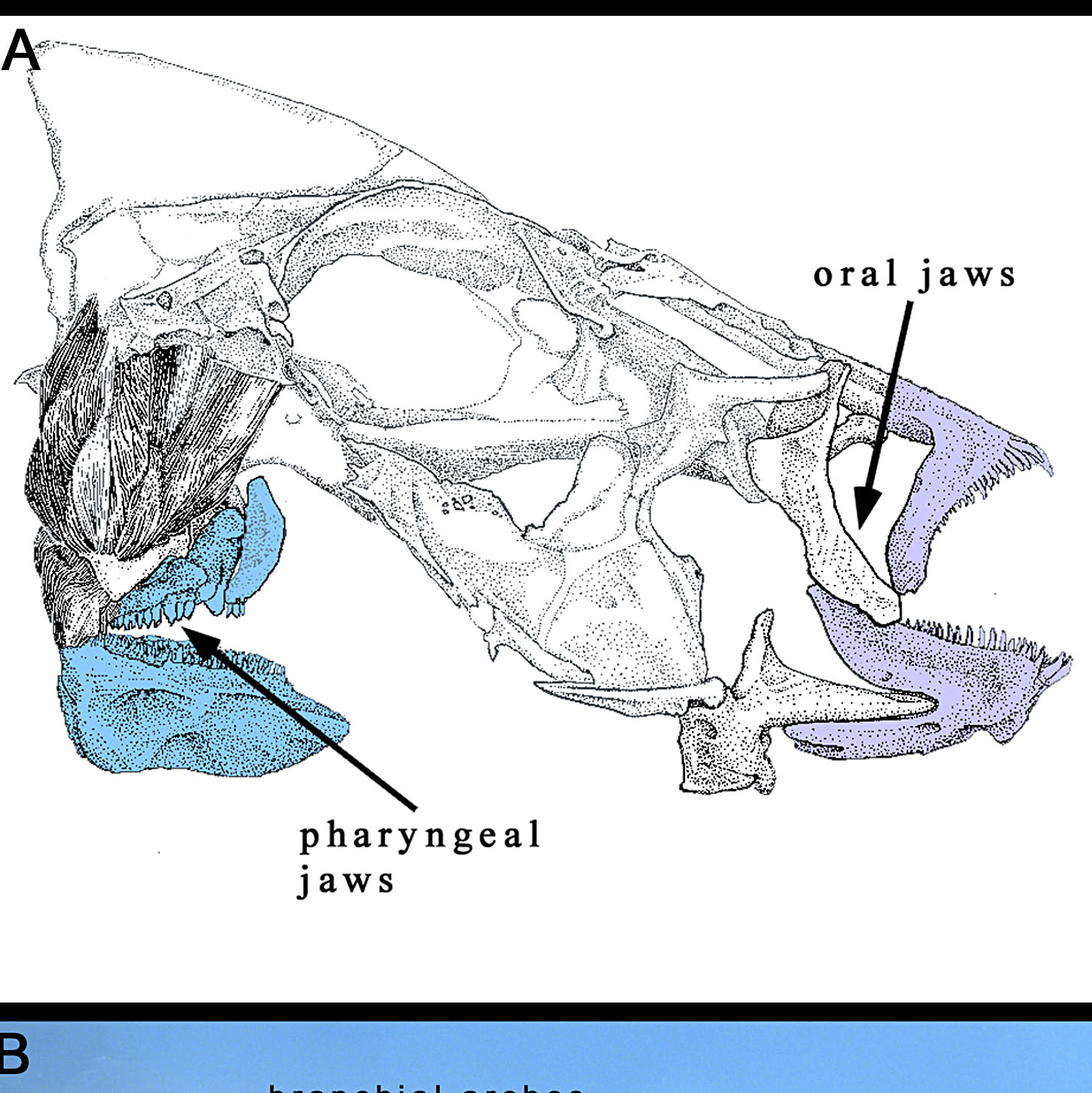
Much of the research into cichlid morphological disparity has long focused on the jaws. Cichlids like many fishes have two pairs of jaws. The oral jaws evolved for prey capture, the pharyngeal jaws evolved for prey processing (Fraser et al., 2009). To some extent the lips might also provide a purpose in some fishes in some fishes to move a surface to extract food further as seen in other fishes (Krings et al., 2023; Cohen et al., 2023).
Figure 1: The jaws of a typical cichlid as featured in: Fraser, G. J., Hulsey, C. D., Bloomquist, R. F., Uyesugi, K., Manley, N. R., & Streelman, J. T. (2009). An ancient gene network is co-opted for teeth on old and new jaws. PLoS biology, 7(2), e1000031.
These two pairs of jaws can be diverse in shape and structure but the teeth further vary between individuals and species.
Cichlids display a very traditional head shape that reflects most other fishes. The oral jaw is often gape limited and evolved to reach forwards to remove or capture a prey item. Realistically no cichlids lack this.
Earlier I mentioned Tropheus being algivores yet it doesn’t seem there is any solid evidence that they exclusively are but we can look at the fishes morphology. They are extremely similar to a fish we know likely consumes a lot of bacteria, algae and similar microbes, discus, Symphysodon (Crampton, 2008). These fishes are not related and this similar morphology is likely due to convergent evolution. Both display a shorter blunter head with strong lips, similar to silver dollars and pacu, Serrasalmidae who some force to break apart at that ‘herbivorous matter’ (Cohen et al., 2023), but they do take it to a more extreme level given they are feeding on plant and fruit matter not algaes and bacteria. Tropheus display some densely packed oral teeth (Richardson-Coy, 2017). This differential oral, mouth morphology could really be due to the differences in what is required to feed on different algaes and in fact, Tropheus are confirmed to specialize in diatoms (Richardson-Coy, 2017), diatoms cling to a surface so much more then the more loose algae/biofilm based diet of Symphysodon (Crampton, 2008) so the more numerous teeth would be more effective. Tropheus oral teeth remind me much more of the jaws of Baryancistrus and similar Loricariids who are scraping algaes of rocks. These teeth might be much more useful for feeding on those Discus, Symphysodon display reduced pharyngeal jaws (Burress, 2016) as this diet might not require the same level of processing as plant matter (hence Serrasalmidae have large robust oral teeth). It seems unclear as to the morphology At the end of the day algivores are diverse, no more needs said but just look at the diversity of algivores in Loricariidae, the queens of algivory/detritivory.
In comparison to Tropheus and Symphysodon would be those that specialize in fishes, Cichla as mentioned before has that large and explosive jaw to reach and consume fish. Fishes are a comparatively more difficult food item to obtain then anything herbivorous so quickly grabbing food much more ahead is beneficial.
The position of jaws is a big clue as to what fishes feed on, terminal mouth’s point forward inferring feeding in front of the fish and is usually associated with carnivory. An inferior mouth points downwards so feeding from the bottom, benthic usually associated with invertebrates and herbivory. Superior mouth’s point upwards and therefore specialize in feeding from the surface. We can clearly see while not extreme Tropheus particularly has an inferior mouth whereas Cichla, being a piscivore is terminal.
While these jaw shapes I mention cover the more traditional cichlids we cannot forget the earth eaters, those fishes who find their food in the substrate, shifting and moving around the sand or silt. While this is generally associated with many Geophagini such as Geophagus, Satanoperca but this clade does include species who are limited in this ability to move substrate e.g. Apistogramma, Mikrogeophagus. Distantly related to these other South American genera is Retroculus, another ‘earth-eater’ (Lopez-Fernandez et al., 2012). The substrate feeding trophic niche seems mostly associated with South American cichlids but it is found in the Rift Valley, although much of this behaviour might be more about breeding sites. Interestingly, it seems these substrate interacting cichlids are more then often mouth-brooders. Substrate digging seems much common in many more Rift Valley clades we keep.
The interesting thing some what I can infer from Burress (2016) is that while jaw shapes might be similar tooth shapes seem divergent between neotropical (fishes from the America’s) and old world (Africa, Asia etc.) regardless of a shared niche. This seems to move away from convergent evolution and multiple solutions to the same problem, the same problem as I noticed being a similar diet.
Tropheus spp. sourced from Wikipedia.
Liem’s Paradox
This is the biggest part of the cichlid story. To put simply Liem’s paradox is the fact that while fishes might display specialist diets and morphology, they still are capable of generalization. This theory is based on the behaviour and morphology of Rift Valley cichlids (Liem, 1980). This I can assume is some factor limiting morphology for further specialization in morphology, and we see these extreme specializations in other clades e.g. Loricariidae, Gymnotiformes and Moryridae. This plays out in the wild where algivores, Lepidophages (scale specialists) and other niches are shown to feed on fishes when given the chance (Golcher-Benavides & Wagner, 2019). Perhaps there is a nutritional reason, but just because a fish will eat something it doesn’t mean they eat it frequently or it is good for them. This plays out in our aquariums, discus Symphysodon will eat smaller tetra yet they are detritivores and Rift Valley algivorous cichlids (Hata et al., 2014).
It is well accepted that fishes feed and act very differently in the aquarium to how they would in the wild but it does occur in the fishes natural habitats (Golcher-Benavides & Wagner, 2019). This theory in fishes is largely based on Rift Valley cichlids, it’s quite clear that Tropheus are much more capable of generalism then Symphysodon. This opportunistic generalism would be limited by certain specialist morphology e.g. the body shape of Symphysodon or mouth shape of angelfish (Pterophyllum). The Symphysodon is limited in it’s ability to feed on fishes and invertebrates but the Pterophyllum is limited in it’s ability to eat algae’s.
This might not be so much of a simple topic with so many taxa. With the focus on Africa, there is so much we seem not to know about the South American clades despite their diversity. Regardless many cichlids while specialist do not seem to take it to the extreme, this might be behind their diversity but also why unlike clades like Siluriforme, they are limited in that morphological disparity.
References:
Aguiar‐Santos, J., deHart, P. A., Pouilly, M., Freitas, C. E., & Siqueira‐Souza, F. K. (2018). Trophic ecology of speckled peacock bass Cichla temensis Humboldt 1821 in the middle Negro River, Amazon, Brazil. Ecology of Freshwater Fish, 27(4), 1076-1086.
Arbour, J. H., & López‐Fernández, H. (2014). Adaptive landscape and functional diversity of Neotropical cichlids: implications for the ecology and evolution of Cichlinae (Cichlidae; Cichliformes). Journal of evolutionary biology, 27(11), 2431-2442.
Burress, E. D. (2016). Ecological diversification associated with the pharyngeal jaw diversity of Neotropical cichlid fishes. Journal of Animal Ecology, 85(1), 302-313.
Cohen, K. E., Lucanus, O., Summers, A. P., & Kolmann, M. A. (2023). Lip service: Histological phenotypes correlate with diet and feeding ecology in herbivorous pacus. The Anatomical Record, 306(2), 326-342.
Crampton, W. G. (2008). Ecology and life history of an Amazon floodplain cichlid: the discus fish Symphysodon (Perciformes: Cichlidae). Neotropical Ichthyology, 6, 599-612.
Fraser, G. J., Hulsey, C. D., Bloomquist, R. F., Uyesugi, K., Manley, N. R., & Streelman, J. T. (2009). An ancient gene network is co-opted for teeth on old and new jaws. PLoS biology, 7(2), e1000031.
Golcher-Benavides, J., & Wagner, C. E. (2019). Playing out Liem’s paradox: opportunistic piscivory across Lake Tanganyikan cichlids. The American Naturalist, 194(2), 260-267.
Hata, H., Tanabe, A. S., Yamamoto, S., Toju, H., Kohda, M., & Hori, M. (2014). Diet disparity among sympatric herbivorous cichlids in the same ecomorphs in Lake Tanganyika: amplicon pyrosequences on algal farms and stomach contents. Bmc Biology, 12, 1-14.
Krings, W., Konn-Vetterlein, D., Hausdorf, B., & Gorb, S. N. (2023). Holding in the stream: convergent evolution of suckermouth structures in Loricariidae (Siluriformes). Frontiers in Zoology, 20(1), 37.
Liem, K. F. (1980). Adaptive significance of intra-and interspecific differences in the feeding repertoires of cichlid fishes. American zoologist, 20(1), 295-314.
Lopez-Fernandez, H., Winemiller, K. O., Montana, C., & Honeycutt, R. L. (2012). Diet-morphology correlations in the radiation of South American geophagine cichlids (Perciformes: Cichlidae: Cichlinae). PLoS One, 7(4), e33997.
Richardson-Coy, R. (2017). Feeding Selectivity of an Algivore (Tropheus brichardi) in Lake Tanganyika. PhD Thesis
Santos, M. E., Lopes, J. F., & Kratochwil, C. F. (2023). East African cichlid fishes. EvoDevo, 14(1), 1.
Wagner, C. E., McIntyre, P. B., Buels, K. S., Gilbert, D. M., & Michel, E. (2009). Diet predicts intestine length in Lake Tanganyika’s cichlid fishes. Functional Ecology, 23(6), 1122-1131.
While my fascination is largely with Loricariids, rasping species are interesting. Unlike the majority of fishes who are limited in what they can eat by the size of their mouth rasping fishes are not and yet ecologically seem very misunderstood. It’s also no lie that I have a soft spot for loaches and have kept quite the diversity of different species.
Sewellia breviventralis
Balitoridae are those dorsio-ventrally flattened loaches from the group Cypriniformes, often referred to the superfamily Cobitoidea. So while Balitoridae are commonly confused with pleco’s, Loricariids they are actually more closely related to carp, barbs and minnows.
Phylogeny of loaches, Cobitoidea from: Wang, Y., Shen, Y., Feng, C., Zhao, K., Song, Z., Zhang, Y., … & He, S. (2016). Mitogenomic perspectives on the origin of Tibetan loaches and their adaptation to high altitude. Scientific reports, 6(1), 29690.
There are many genera we keep under this group in the trade, the most common being Sewellia lineaolata but otherwise followed by a variety of species from the genera Pseudogastromyzon and Gastromyzon. Occasionally other genera appear but as by catch many other Sewellia come into the trade such as SEW001 and S. breviventralis. If you want a rare fish and can do some research, it’s not difficult to find something rare or unusual in this group.
Not all have this extreme suction cup-like morphology, some are much more elongate.
The Habitats
As the name suggests these fishes seem to be highly riverine, rocks and high velocity water. There is little to no botanicals, wood and certainly no plants but this clear water (Randall et al., 2023) is where algae can thrive where plants cannot compete. These fishes thrive in a habitat more similar to some Ancistrus, Chaetostoma or Astroblepus would in South America.
Balitoridae Diets
Balitoridae no doubt shows likely a wide range of diets, there doesn’t seem to be the research on them. While few studies exist of these fishes diets the evidence suggests the genus Pseudogastromyzon feeds largely on algaes, both cyanobacteria and traditional Chlorophytic algaes (Fig 1; Yang & Dudgeon, 2010).
Figure 1: Pseudogastromyzon myersi stomach contents from: Yang, G. Y., & Dudgeon, D. (2010). Dietary variation and food selection by an algivorous loach (Pseudogastromyzon myersi: Balitoridae) in Hong Kong streams. Marine and Freshwater Research, 61(1), 49-56.
Although a later study inferred Pseudogastromyzon myersi feeds on 60-100% of their diet is algaes, with a lot of diversity. In comparison another genus with a similar body morphology, Liniparhomaloptera while feeding on largely algae’s displayed a diet of largely 6-20% invertebrates (Mantel et al., 2004). This is also displayed in Homaloptera sp. which feeds on around 13-14% insects, although the majority of their diet is detritus (Fuadi et al., 2016). As discussed before detritus is a vague classification and could realistically mean anything, it is most likely ‘bacteria’ over waste. A large amount of invertebrates consumed by Homaloptera is ostracods and aquatic larvae, it seems that they feed on minimal algae’s (Nithirojpakdee et al., 2014).
The most unusual thing seems to be this is the only study on the diet of Balitoridae, the majority of studies as with other groups has focused in their phylogenetics and taxonomy (Yang, 2008).
Surprisingly here we have a really good comparison of morphology just externally, compare Gastromyzon, Sewellia etc. with Liniparhomaloptera and immediately their head is much wider. We can some what assume a wider head correlates with a wider jaw and wider jaws, more numerous teeth correlates with a more algivorous diet (Lujan & Armbruster et al., 2012). If we look at the ventral morphology of these fishes one has much wider jaws then the other, Pseudogastromyzon has comparatively wider jaws and general head size then Liniparhomaloptera but is still wider then Homaloptera. The two former species, similar to Sewellia display jaws more similar to Loricariids, plate like jaws with numerous teeth.
Gastromyzon on the left, Sewellia on the right. Image source from: Willis, J., Burt de Perera, T., Newport, C., Poncelet, G., Sturrock, C. J., & Thomas, A. (2019). The structure and function of the sucker systems of hill stream loaches. bioRxiv, 851
There is some obvious jaw diversity in Balitoridae, the mouth of Gastromyzon is much wider then the curved mouth of Sewellia (Willis et al., 2019). I can assume that Gastromyzon is much more similar to Pseudogastromyzon ecologically and regarding diet although they are not closely related (Shao et al., 2020). There is no doubt either species probably feeds on a large amount of algae and other microbial films, but without any ecological records we can only assume.
The shape of the jaw isn’t just about taking up the food but can also be related to being able to extract that food item, while wide, those curved jaws could infer a niche involving extracting microbes from the cracks in rocks and between rocks. Although these cracks, fishers etc. are unlikely to have as many of these microbial films compared to where invertebrates might find refuge. There could be a rugosity aspect as if that habitat has rocks that are naturally bumpy it makes sense to have those more curved jaws then a long jaw which could be unable to access some of these films. A bit like a hoover, you have one part for large flat areas and another to deal with areas that aren’t so flat.
Unlike Loricariids I do not feel looking at their jaws they quite share the exact same niche as they seem to lack the same morphology. Perhaps this is exclusive to siluriformes and there are rasping catfishes in Africa and Asia.
The mouth of Pseudogastromyzon fasciatus as featured in: Chen, J., Chen, Y., Tang, W., Lei, H., Yang, J., & Song, X. (2023). Resolving phylogenetic relationships and taxonomic revision in the Pseudogastromyzon (Cypriniformes, Gastromyzonidae) genus: molecular and morphological evidence for a new genus, Labigastromyzon. Integrative Zoology.
Similar to Loricariids (Krings et al., 2023) it’s is likely not just about the skeletal anatomy that infers diet. The soft tissues at the mouth could play a role as displayed in Chen et al. (2023), there are many ridges on the mouth of the Pseudogastromyzon included these might play a role in sticking to surfaces in their high velocity environment but perhaps when feeding aid in the removal of algaes from a surface.
What should you feed hillstream loaches?
Regardless of the diversity of Balitoridae diets, the majority of their diet is still either detritus and/or algae’s. So I’d build up from that then maybe including a small range of frozen foods for them to forage for.
A brand like Repashy soilent green would be ideal, it is largely algaes with some invertebrates. Alternatively if you can find the algae based New Life Spectrum, AlgaeMax but a product under the same name is higher in fish meals. In the Bag Tropical Fishkeeping UK’s pleco pops would also be great.
It is really tricky to find diets that contain a reasonable volume of algae’s, even if they are listed as algae wafers. Vegetables and cereals do not make up nutritionally for algaes either. At the end of the day if you can’t get these diets then these fishes are not tricky to feed and making your own gel diet is possible, I will maybe write an article about that in future.
References:
Chen, J., Chen, Y., Tang, W., Lei, H., Yang, J., & Song, X. (2023). Resolving phylogenetic relationships and taxonomic revision in the Pseudogastromyzon (Cypriniformes, Gastromyzonidae) genus: molecular and morphological evidence for a new genus, Labigastromyzon. Integrative Zoology.
Fuadi, Z., Naira, K. B., & Hasri, I. 2016. HABITS EATING FISH OF ILI (Homaloptera Sp.) PESTAK RIVER DISTRICT IN CENTRAL ACEH INDONESIA.
Krings, W., Konn-Vetterlein, D., Hausdorf, B., & Gorb, S. N. (2023). Holding in the stream: convergent evolution of suckermouth structures in Loricariidae (Siluriformes). Frontiers in Zoology, 20(1), 37.
Lujan, N. K., & Armbruster, J. W. (2012). Morphological and functional diversity of the mandible in suckermouth armored catfishes (Siluriformes: Loricariidae). Journal of Morphology, 273(1), 24-39.
Mantel, S. K., Salas, M., & Dudgeon, D. (2004). Foodweb structure in a tropical Asian forest stream. Journal of the North American Benthological Society, 23(4), 728-755.
Nithirojpakdee, P., Beamish, F. W. H., & Boonphakdee, T. (2014). Diet diversity among five co-existing fish species in a tropical river: integration of dietary and stable isotope data. Limnology, 15, 99-107.
Randall, Z. S., Somarriba, G. A., Tongnunui, S., & Page, L. M. (2023). Review of the spotted lizard loaches, Pseudohomaloptera (Cypriniformes: Balitoridae) with a re‐description of Pseudohomaloptera sexmaculata and description of a new species from Sumatra. Journal of Fish Biology, 102(1), 225-240.
Shao, L., Lin, Y., Kuang, T., & Zhou, L. (2020). Characterization of the complete mitochondrial genome of Balitora ludongensis (Teleost: Balitoridae) and its phylogenetic analysis. Mitochondrial DNA Part B, 5(3), 2308-2309.
Wang, Y., Shen, Y., Feng, C., Zhao, K., Song, Z., Zhang, Y., … & He, S. (2016). Mitogenomic perspectives on the origin of Tibetan loaches and their adaptation to high altitude. Scientific reports, 6(1), 29690.
Willis, J., Burt de Perera, T., Newport, C., Poncelet, G., Sturrock, C. J., & Thomas, A. (2019). The structure and function of the sucker systems of hill stream loaches. bioRxiv, 851592.
Yang, Y. (2008). The ecology of a herbivorous fish (Pseudogastromyzon myersi: balitoridae) and its influence on benthic algal dynamics in four HongKong streams. HKU Theses Online (HKUTO).
Yang, G. Y., & Dudgeon, D. (2010). Dietary variation and food selection by an algivorous loach (Pseudogastromyzon myersi: Balitoridae) in Hong Kong streams. Marine and Freshwater Research, 61(1), 49-56.
With a whole section designated to Loricariidae, I haven’t actually done a beginners guide to the group. This website largely isn’t designed for beginners but Loricariid’s are some of the most misunderstood group of fishes.
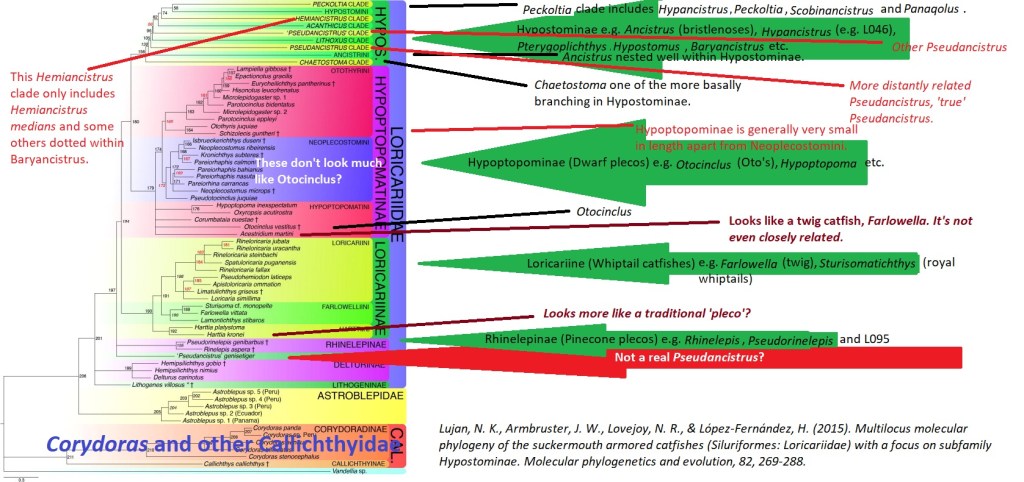
These two common names have no certain definitions, the majority of their use is a pick and mix that varies between the user. It is even more confusing that some loaches are referred to as pleco’s. All common names are equally as valid as each other. I find it easier to refer to the whole of Loricariidae as plecos, why? Figure 1 explains this situation. By excluding the subfamily Loricariinae (whiptail catfishes), you exclude Loricariichthys of which Plecostomus was synonymized with. If you exclude Hypoptopominae (Otocinclus and relatives) then Neoplecostomini and Neoplecostomus are excluded. Ancistrus, commonly known as bristlenose’s places right in the middle of Hypostominae, traditional plecos but Ancistrus also includes the medusa pleco, Ancistrus ranunculus. Then outside of all of these groups is Rhinelepinae, so that includes the pineapple pleco’s, and on it’s own Pseudancistrus genisetiger, so none of those are plecos then?
What would solve the common name issue? Simply not using them. Sadly with Loricariids you can’t avoid scientific names as many species do lack common names or share them.
Figure 1: Annotated phylogeny of Loricariidae from Roxo, F. F., Ochoa, L. E., Sabaj, M. H., Lujan, N. K., Covain, R., Silva, G. S., … & Oliveira, C. (2019). Phylogenomic reappraisal of the Neotropical catfish family Loricariidae (Teleostei: Siluriformes) using ultraconserved elements. Molecular phylogenetics and evolution, 135, 148-165.
What is the L number system?
The L number system is actually quite simple, it is a hobby made system originating from the German Magazine, DATZ. It simply designates an L number to a variant or species. It is commonly stated that undescribed species are given L numbers, this is partially true but there are many species who were described decades or over 100 years before given their L number such as the sailfin/common/leopard pleco, Pterygoplichthys gibbiceps who was described by Kner in 1854.
I am not entirely convinced the L number system is easy to use, the order of the numbers doesn’t infer anything regarding the fishes care or lineages. Multiple species can share an L number e.g. Baryancistrus demantoides and Hemiancistrus subvirdis are both L200 also known as green phantoms. Hemiancistrus subvirdis is likely the same species as L128 although that is a topic for another day. One species can have multiple L numbers, which don’t always describe populations or all of the morphological variation of the species e.g. Baryancistrus xanthellus has 4 L numbers.
To top it off there are fake L numbers, L600 for example described Pseudacanthicus leopardus who already has the L numbers, L114 and L427. The L number system is only up to around 530 species so far. The letters added to L numbers aren’t a part of the L number system and increase confusion. Sometimes species are given by the hobby the L number of an entirely different species such as L144 which doesn’t even exist in the hobby and hasn’t done for decades or L056 for Parancistrus aurantiacus when actually that L number refers to an undescribed brown Pseudancistrus.
There is additionally the LDA number system which does overlap slightly but isn’t so expansive.
Introduction
Loricariidae, pleco’s are the largest family within the order of fishes known as Siluriforme also known as catfishes or welse, representing 1050 currently described species (Fricke et al., 2024). This group is exclusive to South and Central American freshwaters although has invasive populations in many continents.
Panaque nirolineatus from Maidenhead Aquatics at Ascot.
The family is identified by a downwards (ventrally) facing oral disc shaped mouth, in some species this is more of a suction cup whereas others they cannot attach to surfaces well or at all. This trait isn’t exclusive to Loricariids but it is not quite the same in other groups. Further more, Loricariids are defined by having a body covered in bony scutes, more scientifically known as dermal plating, not scales as catfishes lack scales.
Baryancistrus chrysolomus.
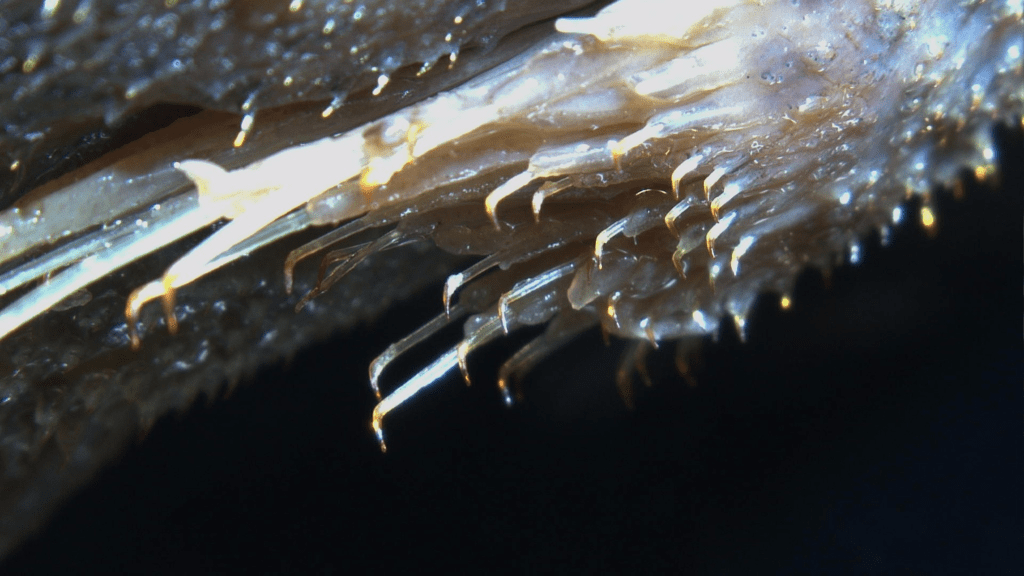
Not does Loricariidae just have dermal plating but they have spines known as odontodes, external teeth (Fig 2). Sometimes these are sexually dimorphic but not always, they can also be shed seasonally.
Figure 2: Odontodes found on I believe Peckoltia sabaji.
Loricariids realistically are one of the most morphologically diverse clades of fishes.
This diversity means that as a group, Loricariids are really difficult if impossible to generalize, research is paramount for this group.
The size of plecos
As one of the most diverse groups of fishes their size varies vastly, 0.2-0.8cm in Parotocinclus halbothi (Lehmann et al., 2014) to 100cm SL possibly in Acanthicus adonis. There is a wide diversity of sizes within many groups so there is no shortage of smaller and larger species. At the end of the article I will recommend reliable websites, there is frequent misleading information about the adult size of many species.
It is important to recognize reliable websites will use standard length, from the head to the base of the caudal/tail fin. That caudal/tail fin will be excluded as these can vary in length. I mention this as many people will not consider this measurement and forget their fish grows much bigger then they would originally consider. This is explained in detail in this article.
There are a few myths regarding Loricariid diets I will summerise:
Pleco’s are largely carnivores, there are plenty of papers discussing their diets and while I wont cite them all Lujan et al. (2015) summerises it well.
Pleco’s become carnivorous with age, there isn’t any studies regarding change in diet as the fishes age. Unlike most fishes, the majority of Loricariids break down their food before it enters their mouth, so the size of the fish doesn’t limit their food item, so not gape limited. This means unlike many other fishes their food item doesn’t need to change with size.
Pleco’s clean a tank, while the majority are algivores and detritivores (Lujan et al., 2015) there is a wide diversity in niche partitioning (Lujan et al., 2011) and therefore those algivore’s specialize in certain algae’s. These algae’s seem not to be those that are an issue in the aquarium. Given their lifespan and waste production, they could be an expensive solution to high nutrients.
Pleco’s don’t eat cyanobacteria, they actually don’t just eat those that are pests in the aquarium, in the wild they feed on cyanobacteria (Valencia & Zamudio ,2007).
Always check the ingredients as some diets that claim to contain algae’s might contain anything from none to 5%.
In simple NO, they do not need wood. The only species that utilize wood are in the genera Panaque, Panaqolus, Hypostomus cochliodon group, Pseudoqolus and perhaps Lasiancistrus heteracanthicus. These groups all share unique spoon shaped teeth they can gouge into wood and if found among wood, wood is found in their gut (Lujan et al., 2017). No other Loricariid has wood in their gut, I’ve scoured gut records but they simply don’t have the jaws or teeth to gouge into wood. There have been many studies to confirm these fishes do not eat/digest the wood (Watts et al., 2021; German, 2009), instead they are just evolved to feed on biofilms, like other species from where other species cannot access, within wood (Lujan et al., 2011).
Peckoltia compta
What parameters do pleco’s need?
While the general idea is that Loricariids and in general anything South America requires soft, low conductivity and acidic water there is a wide diversity of parameters. Some species are found in lower temperatures while other much higher, 28c or higher (Collins et al., 2015; Urbano‐Bonilla & Ballen et al., 2021). Other misconceptions are that South American habitats have a lot of leaf litter and is black water, this is completely untrue, there are many different habitat types (Bogotá-Gregory et al., 2020). In general the majority of Loricariidae are rheophilic and would benefit from a current within the aquarium although there is some diversity (Krings et al., 2023). In general any current within the aquarium is a lot weaker then any of the weaker streams in their wild range.
Planet Catfish has really accessible information to identify parameters before looking into the scientific literature.
What decor do plecos require?
This is largely the only consistent aspect of Loricariids. If anyone has kept Loricariids they will know how much they like cracks, crevices, hiding spaces, rocks, branches and in general cover. There are many caves and tunnels on the market designed for the preferences of a variety of species. I recommend stacking up wood or rocks in a careful way so nothing falls but this will create many more caves.
Tankmates
This will always be based on experience and understanding of the fishes. It’s important to recognize a few key things about Loricariids.
Loricariids do not often feed rapidly but even if they do it can take them minutes to an hour to reach food. Fast feeding fishes such as most cichlids, loaches, tetra, livebearers and goldfish particularly in large numbers are a bad idea.
Their temperature might not overlap.
They will need a current, some more then others which this means they wont work with fishes like long finned Betta splendens.
While some Loricariids feed on food items that they wouldn’t naturally it doesn’t mean it is good for them. Bloat can happen in some genera more then others. So I do not recommend keeping other fishes with Loricariids who you plan on feeding anything like beefheart.
Hardness, conductivity etc. We don’t actually know the KH or GH of the water many of these fishes come from, usually we have conductivity and pH records for many waters. Ideally these fishes are ill-suited regardless with Rift Valley cichlids, generally the biggest issue is above, those cichlids feed way too rapidly for any Loricariid.
Collins, R. A., Ribeiro, E. D., Machado, V. N., Hrbek, T., & Farias, I. P. (2015). A preliminary inventory of the catfishes of the lower Rio Nhamundá, Brazil (Ostariophysi, Siluriformes). Biodiversity Data Journal, (3).
Bogotá-Gregory, J. D., Lima, F. C., Correa, S. B., Silva-Oliveira, C., Jenkins, D. G., Ribeiro, F. R., … & Crampton, W. G. (2020). Biogeochemical water type influences community composition, species richness, and biomass in megadiverse Amazonian fish assemblages. Scientific Reports, 10(1), 15349.
German, D. P. (2009). Inside the guts of wood-eating catfishes: can they digest wood?. Journal of Comparative Physiology B, 179, 1011-1023.
Krings, W., Konn-Vetterlein, D., Hausdorf, B., & Gorb, S. N. (2023). Holding in the stream: convergent evolution of suckermouth structures in Loricariidae (Siluriformes). Frontiers in Zoology, 20(1), 37.
Lehmann, P. A., Lazzarotto, H., & Reis, R. E. (2014). Parotocinclus halbothi, a new species of small armored catfish (Loricariidae: Hypoptopomatinae), from the Trombetas and Marowijne River basins, in Brazil and Suriname. Neotropical Ichthyology, 12, 27-33.
Lujan, N. K., Cramer, C. A., Covain, R., Fisch-Muller, S., & López-Fernández, H. (2017). Multilocus molecular phylogeny of the ornamental wood-eating catfishes (Siluriformes, Loricariidae, Panaqolus and Panaque) reveals undescribed diversity and parapatric clades. Molecular phylogenetics and evolution, 109, 321-336.
Lujan, N. K., German, D. P., & Winemiller, K. O. (2011). Do wood‐grazing fishes partition their niche?: morphological and isotopic evidence for trophic segregation in Neotropical Loricariidae. Functional Ecology, 25(6), 1327-1338.
Lujan, N. K., Winemiller, K. O., & Armbruster, J. W. (2012). Trophic diversity in the evolution and community assembly of loricariid catfishes. BMC Evolutionary Biology, 12, 1-13.
Roxo, F. F., Ochoa, L. E., Sabaj, M. H., Lujan, N. K., Covain, R., Silva, G. S., … & Oliveira, C. (2019). Phylogenomic reappraisal of the Neotropical catfish family Loricariidae (Teleostei: Siluriformes) using ultraconserved elements. Molecular phylogenetics and evolution, 135, 148-165.
Urbano‐Bonilla, A., & Ballen, G. A. (2021). A new species of Chaetostoma (Siluriformes: Loricariidae) from the Orinoco basin with comments on Amazonian species of the genus in Colombia. Journal of Fish Biology, 98(4), 1091-1104.
Valencia, César Román, and Héctor Zamudio. 2007. Dieta y reproducción de Lasiancistrus caucanus (Pisces: Loricariidae) en la cuenca del río La Vieja, Alto Cauca, Colombia. Revista del Museo Argentino de Ciencias Naturales nueva serie 9(2): 95-101.
Watts, J. E., McDonald, R. C., & Schreier, H. J. (2021). Wood degradation by Panaque nigrolineatus, a neotropical catfish: diversity and activity of gastrointestinal tract lignocellulolytic and nitrogen fixing communities. In Advances in Botanical Research (Vol. 99, pp. 209-238). Academic Press.
Bloodworm has been a food that has been discouraged against for around 10 or more years. There has never been any doubt that many fishes feed on Chironomatid larvae in the wild (Delariva & Agostinho, 2001; Valtierra-Vega & Schmitter-Soto, 2000), even discus feed on them (Crampton, 2008). Chironomatid larvae being the more scientific name for bloodworm. You might have even seen these larvae in your pond, buckets or similar.
Bloodworms as featured in: Paice, R. (2016). Assessment of mosquito larvicide impacts on aquatic invertebrates in the Vasse-Wonnerup Wetland System.
So why are bloodworms so feared?
I don’t think there is a true origin to this, it is still one of the most common live and frozen foods used, it’s cheap and easy to find. Maybe there are still some aspects of when tubifex was feared due to sewage contamination.
Bloodworm being the larvae of an insect does have a casing, known as an exoskeleton formed of chitin. This can be difficult to digest and for some fishes impossible, although many species are possibly sold as bloodworm it’s likely there is some diversity. There are reports of constipation and similar caused by bloodworm, although bloating is tricky and can have many causes from bacterial infections to a food item not being processed.
Bloodworm is not the entire diet of any fishes I’ve researched and when fed as an entire diet it results in liver enlargement and and poorer body condition (Žák et al., 2022), these results are similar to Hao et al. (2021) where removing to a formulated diet increases general health of the fish.
I chose this topic to find reasons that I am wrong, that bloodworm is much better then I thought it was. It doesn’t seem bad for fattening up fishes but does seem to come with a variety of problems, as a result I think I will continue to recommend Tubifex.
The real worm to try
Tubifex is a relatively common live, frozen and freeze dried food available. Unlike bloodworm it’s an annelid, true worm and is fully aquatic. It lacks the thick chitin casing of bloodworms, making them much more easy to digest. To cultivate they are less tricky given their lifestyle but do need a constant flow of freshwater.
Unlike bloodworm, Tubifex was vilified for years, this amazing genus is extremely adaptable. This adaptability has meant that Tubifex can be found in the most hostile and polluted habitats and at one time this is where they were collected for the aquarium trade. Due to being collected from very polluted habitats it was not the most ideal live food to avoid pollution. Frozen will be another story as most are gamma irradiated, killing most bacteria on them. But for many years Tubifex is farmed in a clean and sustainable matter.
Unlike bloodworm Tubifex is very high in nutrition (Herawati et al., 2016) and has shown to increase growth when mixed with a prepared diet (Alam et al., 2021) even when compared to other live foods (Mellisa et al., 2018).
Human health
Bloodworm is a known allergen leading to asthma (Wu et al., 2005; Nandi et al., 2014). As someone who is not a medical practitioner I cannot offer much advice, allergies can occur at any time and it is good to be cautious. When dealing with bloodworm, handling particularly wearing disposable gloves can be a good idea, using a tub to defrost it in. It does beg the question given it is quite a common allergen whether to use it at all? In my experience I have met very few people who are allergic to bloodworm.
References:
Alam, M. A., Khan, M. A., Sarower-e-Mahfuj, M. D., Ara, Y., Parvez, I., & Amin, M. N. (2021). A model for tubificid worm (Tubifex tubifex) production and its effect on growth of three selected ornamental fish. Bangladesh Journal of Fisheries, 33(2), 205-214.
Crampton, W. G. (2008). Ecology and life history of an Amazon floodplain cichlid: the discus fish Symphysodon (Perciformes: Cichlidae). Neotropical Ichthyology, 6, 599-612.
Delariva, R. L., & Agostinho, A. A. (2001). Relationship between morphology and diets of six neotropical loricariids. Journal of Fish biology, 58(3), 832-847.
Hao, Q., Teame, T., Wu, X., Ding, Q., Ran, C., Yang, Y., … & Zhou, Z. (2021). Influence of diet shift from bloodworm to formulated feed on growth performance, gut microbiota structure and function in early juvenile stages of hybrid sturgeon (Acipenser baerii× Acipenser schrenckii). Aquaculture, 533, 736165.
Herawati, V. E., Nugroho, R. A., Hutabarat, J., & Karnaradjasa, O. (2016). Profile of amino acids, fatty acids, proximate composition and growth performance of Tubifex tubifex culture with different animal wastes and probiotic bacteria. Aquaculture, Aquarium, Conservation & Legislation, 9(3), 614-622.
Mellisa, S., Rahimi, S. A. E., & Umiati, U. (2018). The effect of different live feeds on the growth and survival of comet goldfish Carrasius auratus auratu larvae. In IOP Conference Series: Earth and Environmental Science (Vol. 216, No. 1, p. 012025). IOP Publishing.
Nandi, S., Aditya, G., Chowdhury, I., Das, A., & Saha, G. K. (2014). Chironomid midges as allergens: evidence from two species from West Bengal, Kolkata, India. Indian Journal of Medical Research, 139(6), 921-926.
Valtierra-Vega, M. T., & Schmitter-Soto, J. J. (2000). Feeding habits of cichlid species (Perciformes: Cichlidae) in Caobas lake, Quintana Roo, Mexico. Revista de Biologia Tropical, 48(2-3), 503-508.
Wu, K. C., Räsänen, K., & Hudson, T. J. (2005). Fishing for allergens: bloodworm-induced asthma. Allergy, Asthma & Clinical Immunology, 1, 1-2.
Žák, J., Roy, K., Dyková, I., Mráz, J., & Reichard, M. (2022). Starter feed for carnivorous species as a practical replacement of bloodworms for a vertebrate model organism in ageing, the turquoise killifish Nothobranchius furzeri. Journal of Fish Biology, 100(4), 894-908.
Fake news was the word of the year for 2017, we are often bombarded by information and it can be very difficult to fact check. Sometimes we just don’t have time to check the information we are given is correct, and other times we are relying on the credential of the writer. The aquarium hobby is no exception with so many different websites and social media platforms all arguing for space and more importantly your trust.
As distrust in authority increases there has never been so much division and this leads into the influences of certain media. Unlike the general hobby there is a strong influence of science, whether the website is actually scientific or not.
This is best discussed in terms of two of the biggest myths that have arisen in the last few years.
How Epistylis ruled the world.
For many years the protozoan whitespot, Ichthyophthirius multifiliis was widely considered the most common parasite on fishes, it is well studied in the scientific literature (Francis-Floyd et al., 2016, 2023) followed by velvet, Oodinium and Piscinoodiniasis. Disease is tricky, unlike identifying animals we don’t necessary have the tools as a hobby for an accurate diagnosis, there are thousands of species to separate.
Realistically these two pathogens are one of many that can appear as spots on the fishes body, generally most treatments will cover them. Although there is suggestion that the formalin based treatments are less effective as I discuss here, maybe this has lead to why people come to think they are not dealing with white spot? Because shouldn’t white spot treatments work for I. multifiliis? In addition there is proven acquired immunity to I. multifiliis (Teixeira Alves and Taylor, 2020) but it doesn’t mean that it is unseen. In fact I have noticed a reduced number personally and online of those spotted parasitic cysts, but that is just experience. To add to the confusion not all of these parasites do appear as spots (Sudhagar et al., 2022; Fig 1). I. multifilliis and Piscinoodiniasis is known to target the gills of fishes.
An infection of Piscinoodinium as featured in: Sudhagar, A., Sundar Raj, N., Mohandas, S. P., Serin, S., Sibi, K. K., Sanil, N. K., & Raja Swaminathan, T. (2022). Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius). Pathogens, 11(11), 1350.
This confusion isn’t aided by nematode cysts looking incredibly similar to I. multifilliis and that used to be quite a common misdiagnosis. Going back to the difficulties in identification, microscopes are very important here as many of these pathogens are really only different to look at under the microscope. Using a microscope is one thing but identifying what you are looking at is another, the koi world seems to have this sorted with pathologists available but it seems to have been a reduced talent within the aquarium hobby. There are very few useful books on pathology still available, most if all will be second hand. Microscopes are really only useful for what can be seen so bacteria and even more viruses are another ball game, requiring expensive technology to identify the pathogen.
So, the story is set for Epistylis. Why Epistylis? We will never know but it is likely a random page that came up and sometimes names stick. Unlike Ichthyophthirius multifiliis, Epistylis is easy to remember. I don’t think the why matters but more that it is there, it’s difficult to work out what website started it.
Figure 2: Whitespot, Ichthyophthirius multifiliis as featured in: Martins, M. L., Cardoso, L., Marchiori, N., & Benites de Pádua, S. (2015). Protozoan infections in farmed fish from Brazil: diagnosis and pathogenesis. Revista Brasileira de Parasitologia Veterinária, 24, 1-20.
It might be that fact checking takes time and we should trust what we read to be true. but Ichthyophthirius multifiliis looks nothing alike Epistylis in any way that can be confused as you can clearly see in figure 2 and 3.
Figure 3: Epistylis as featured in: Martins, M. L., Cardoso, L., Marchiori, N., & Benites de Pádua, S. (2015). Protozoan infections in farmed fish from Brazil: diagnosis and pathogenesis. Revista Brasileira de Parasitologia Veterinária, 24, 1-20.
What is more clear is that Epistylis is almost always present and not always as a pathogen, it can also be asymptomatic (Ksepka et al., 2021). But if symptomatic it is expressed as more of a plaque that could be confused with some of the herpes viruses (Fig 3).
So why are they confused? It has to do with this graph (Fig 4). Firstly ich is referring to Ichthyophthirius multifiliis, that we more then often call whitespot in the UK. No citations are provided so this is not backed up with the identification of Epistylis. So some will say the science is wrong, the issue is that genera are described with set features and if those features don’t match then that is not that species, it would be another. That’s how scientific descriptions work. Epistylis doesn’t display itself as distinct round spots whether rough sized or not (Ksepka et al., 2021; Wu et al., 2021; Valladao et al., 2015). If it was so common and lethal there would be more literature when in reality there isn’t that much comparatively. In my experience as well white spot can kill a fish rapidly, and due to the effect on the gills it is quite taxing on the fish (Martins et al., 2015).
So why this whole essay? To discuss misinformation I must first state why this myth isn’t true. Interestingly it is a brilliant story regarding understanding critical analysis.
How to identify a reliable website:
Ability to cite their sources, ideally papers if they are scientists.
Using standard length, just because there are no reliable measurements of total size including the caudal/tail fin.
Avoiding plagiarism, if a scientist this is a key issue.
References:
Francis-Floyd, R., Yanong, R., & Pouder, D. (2023). Ichthyophthirius multifiliis (white spot) infections in fish.
Francis-Floyd, R., Yanong, R., & Pouder, D. (2016). Ichthyophthirius multifiliis (White Spot) Infections in Fish: CIR920/FA006, rev. 12/2016. EDIS, 2016(10).
Ksepka, S. P., & Bullard, S. A. (2021). Morphology, phylogenetics and pathology of “red sore disease”(coinfection by Epistylis cf. wuhanensis and Aeromonas hydrophila) on sportfishes from reservoirs in the South‐Eastern United States. Journal of Fish Diseases, 44(5), 541-551.
Martins, M. L., Cardoso, L., Marchiori, N., & Benites de Pádua, S. (2015). Protozoan infections in farmed fish from Brazil: diagnosis and pathogenesis. Revista Brasileira de Parasitologia Veterinária, 24, 1-20.
Sudhagar, A., Sundar Raj, N., Mohandas, S. P., Serin, S., Sibi, K. K., Sanil, N. K., & Raja Swaminathan, T. (2022). Outbreak of Parasitic Dinoflagellate Piscinoodinium sp. Infection in an Endangered Fish from India: Arulius Barb (Dawkinsia arulius). Pathogens, 11(11), 1350.
Teixeira Alves, M., & Taylor, N. G. (2020). Models suggest pathogen risks to wild fish can be mitigated by acquired immunity in freshwater aquaculture systems. Scientific Reports, 10(1), 7513.
Valladao, G. M. R., Levy-Pereira, N., Viadanna, P. H. D. O., Gallani, S. U., Farias, T. H. V., & Pilarski, F. (2015). Haematology and histopathology of Nile tilapia parasitised by Epistylis sp., an emerging pathogen in South America. Bulletin of the European Association of Fish Pathologists, 35(1), 14-20.
Wang, Z., Zhou, T., Guo, Q., & Gu, Z. (2017). Description of a new freshwater ciliate Epistylis wuhanensis n. sp.(Ciliophora, Peritrichia) from China, with a focus on phylogenetic relationships within family Epistylididae. Journal of Eukaryotic Microbiology, 64(3), 394-406.
Wu, T., Li, Y., Zhang, T., Hou, J., Mu, C., Warren, A., & Lu, B. (2021). Morphology and molecular phylogeny of three Epistylis species found in freshwater habitats in China, including the description of E. foissneri n. sp.(Ciliophora, Peritrichia). European Journal of Protistology, 78, 125767.
Loricariidae, also known under the common names L numbers, whiptail catfishes and pleco’s are popular fishes within the aquarium trade. Many people will come across the problem in their new fish where they will not eat and in fact they might never eat.
Worming
The first sign might be that the fish might have a concaved stomach and the first solution will be to worm the fish. I doubt this is the usual cause of the concave stomach in Loricariid’s but it is worth crossing out, Loricariids do quite frequently have parasitic worm’s (usually nematodes rather then Annelid’s) in the wild and these will maintain at a low level (Borges et al., 2018). If a fish is stressed such as from import this parasite load can become much higher then a healthy level. So there is logic in worming fishes when they arrive and most stores do this. Most wormers cover different internal parasites but the most common would be containing praziquantel, levamisole and flubendazole (do not use with stingrays). I would personally advise definitely not using more then one as they do all have side effects. Generally wormers need to be repeated after a week to cover the parasites lifecycle.
Panaqolus aff. maccus
The importance of getting the fish feeding.
While a rounded and healthy diet is important for wild caught fishes particularly they do need to feed. It is quite a large jump for many from a wild diet to a captive diet and many might not even identify it as food.
More importantly it is possible that the gut flora, microbes will start to decline in number while they are not eating and for shipping this is useful but not for keeping the fish. One possible thing that could help this is rather then adding them to a clean quarantine tank is to one where other similar species have lived, there is likely a benefit from those fishes waste in rebuilding that gut flora lost after shipping.
What should I feed my fish?
First identify what they eat, so their natural diet. While most diets contain the steryotypical fish/insect/krill meal, cereal, vegetable and minimal algae diets this is no issue in the short term but many wont touch these diets at first. So regardless of long term them being vastly different from their natural diet and homogenous whether you have a Trophius, Loricariid or angelfish they actually are the same (Vucko et al., 2017); they can also be unhelpful.
This is important as feeding an incorrect diet can lead to bloat and other issues, it has been commonly noted when Hypancistrus are fed a wholly carnivorous diet.
Carnivorous species
This is only for true carnivores but aspects of this can be fed to others in small numbers with care, avoid it with some of the more extreme algivores such as Ancistrus, Chaetostoma, Baryancistrus etc.
Mussels and prawns are very good for getting a fish feeding at first but the issue with these two food items at high in thiaminase and therefore degrade thiamin, vitamin b1.
Generally for this reason I’d advise a range of frozen foods and for some larger species earthworms might not be a bad choice.
Nannopotopoma sp. ‘Peru/robocop’ at Maidenhead Aquatics at Ascot
Algivores and Detritivores
This includes most Loricariids that people keep to some degree but the specialities within their diet are best looking at later.
These are definitely the most tricky to get feeding at first and I often give a range of options even at the same time. Generally I’d offer that dry/gel diet once or twice a day and vegetables replaced every 12-24 hours depending on how quickly they are braking down.
For dry/gel diets I’d offer certainly Repashy soilent green if possible as I’ve never had a fish fail to give it a go. Later on I’d bulk it out with other ingredients such as algal powders, you could do similar with other gel diets but I can’t say fishes are going to take up them as well. At the end of the day whatever they are eating in the short term is worth it. Remember vegetables and similar are more treats as do not even closely replicate their wild diets.
Vegetables and other easy food items you can leave in for the fishes:
Courgette
Reasonable in nutrition, is willingly eaten by many fishes but they might select either the flesh or skin over the other.
Cucumber
While often declared as low nutrients due to water content they do contain minerals and other compounds that have nutritional value.
Mushrooms (Edible species from supermarket)
Could be part of a staple diet for Panaqolus, Panaque and Hypostomus cochliodon group as they do feed on fungi in the wild (Lujan et al., 2011). It is difficult to say the nutrition levels for these fishes as many might be able to digest more so then nutritional estimates for humans. So far mushrooms are shown to increase weight gain opposed to traditional diets (Zakaria et al., 2021; Dawood et al., 2010), a potential prebiotic (Chandra & Qureshi, 2023) and other potential benefits (Sánchez-Velázquez et al., 2014)
Sweet Potatoes
These doesn’t need to be blanched and I am not convinced by their digestibility for Loricariids (Omoregie et al., 2009) but if they can get the fish feeding that is what matters.
Further on I find whether fishes feed on these more hit and miss.
Green beans
The common bristlenose, Ancistrus sp. is meant to be a big fan of this. Nutrition doesn’t need to be debated but as a plant would be more of a treat after acclimation.
Bell peppers
I don’t think it entirely matters whether the pepper is red, yellow or green but the sugar and nutrition levels will vary.
Pumpkins and other squashes
I find very hit or miss but never blanched them. They can break down very quickly producing a film over the fruit. I would say they are much more similar to courgette.
Later on and narrowing down the diet
While whatever they will eat is generally the best rule as they are acclimatizing over the first few weeks and months. Afterwards I would look to narrowing down their diet to what they would feed on in the wild as in the articles mentioned earlier on.
Is the setup right?
This sometimes get’s forgotten but a major part of why a fish might not be feeding could be they are not getting to the food. Loricariids are slow to feed, some might take hours even without lights to feed and this can make some tankmates ill-suited. Some tankmates might work better where if needed you can remove them to another tank so that is worth considering particularly for many cichlids, many shoaling species in very high numbers or quite a few live numbers.
Planiloricaria cryptodon at Maidenhead Aquatics at Ascot.
References:
Borges, W. F., de Oliveira, M. S. B., Santos, G. G., & Tavares-Dias, M. (2018). Parasites in Loricariidae from Brazil: checklist and new records for fish from the Brazilian Amazon. Acta Scientiarum. Biological Sciences, 40, 1-9.
Chandra, O. P., & Qureshi, Y. (2023). Importance of mushroom supplementation as a prebiotic amalgamation in fed diet of improvement of weight gain (WG) in Nile Tilapia,(Oreochromis niloticus). Journal of Pharmaceutical Negative Results, 1681-1687.
Dawood, M. A., Eweedah, N. M., El-Sharawy, M. E., Awad, S. S., Van Doan, H., & Paray, B. A. (2020). Dietary white button mushroom improved the growth, immunity, antioxidative status and resistance against heat stress in Nile tilapia (Oreochromis niloticus). Aquaculture, 523, 735229.
Lujan, N. K., German, D. P., & Winemiller, K. O. (2011). Do wood‐grazing fishes partition their niche?: morphological and isotopic evidence for trophic segregation in Neotropical Loricariidae. Functional Ecology, 25(6), 1327-1338.
Omoregie, E., Igoche, L., Ojobe, T. O., Absalom, K. V., & Onusiriuka, B. C. (2009). Effect of varying levels of sweet potato (Ipomea Batatas) peels on growth, feed utilization and some biochemical responses of the cichlid (Oreochromis Niloticus). African Journal of Food, Agriculture, Nutrition and Development, 9(2), 700-712.
Sánchez-Velázquez, J., Peña-Herrejón, G. A., & Aguirre-Becerra, H. (2024). Fish Responses to Alternative Feeding Ingredients under Abiotic Chronic Stress. Animals, 14(5), 765.
Vucko, M. J., Cole, A. J., Moorhead, J. A., Pit, J., & de Nys, R. (2017). The freshwater macroalga Oedogonium intermedium can meet the nutritional requirements of the herbivorous fish Ancistrus cirrhosus. Algal research, 27, 21-31.
Zakaria, Z., Abd Rasib, N. A., & Tompang, M. F. (2021). Spent mushroom substrate based fish feed affects the growth of catfish (Clarias gariepinus). In IOP Conference Series: Earth and Environmental Science (Vol. 765, No. 1, p. 012082). IOP Publishing.
Corydoras is a genus of Catfishes in the family Callichthyidae. Callichthyidae includes: Callichthyinae (Callichthys, Lepthoplosternum, Hoplosternum, Megalechis and Dianema); Corydoralinae which is the focus of this article and revision. Figure 1 explains the topography (family tree format) of this group but bare in mind it doesn’t reflect the current knowledge for Corydoralinae.
Figure 1: Phylogeny of Callichthyidae. Bare in mind the topography (shape) of Corydoras doesn’t match our current understanding. As featured in: Shimabukuro-Dias, C. K., Oliveira, C., Reis, R. E., & Foresti, F. (2004). Molecular phylogeny of the armored catfish family Callichthyidae (Ostariophysi, Siluriformes). Molecular Phylogenetics and Evolution, 32(1), 152-163.
Previously many genera were inferred for members of what was known for a long time as Corydoras but over time these were synonymized finalizing with Britto (2003) which resulted in Brochis no longer being valid. To put it simply as there were many reasons behind but also to have Corydoras as the genus it was then Brochis would have to be Corydoras.
Figure 2: Phylogeny of Corydoralinae featured in: Alexandrou, M. A., Oliveira, C., Maillard, M., McGill, R. A., Newton, J., Creer, S., & Taylor, M. I. (2011). Competition and phylogeny determine community structure in Müllerian co-mimics. Nature, 469(7328), 84-88.
The history is a bit messy. We have known for a long time Corydoras has had a lineage system, this was explained clearly in Alexandrou et al (2011). This phylogeny shows how embedded Brochis is inside of Corydoras and therefore would not easily be valid as a genus.
The issue of type species
So while many of the ‘Corydoras‘ kept in the aquarium trade are not Corydoras, the type, C. geoffroy places in lineage 1. These would be the only true Corydoras, basically this is the reference species in a way.
Some might note what was known as Aspidoradini, a tribe separate to what was Corydoras branches out in the middle of Corydoras, more problems. It’s really messy.
What are the new genera?
Corydoras: Used to represent lineage 1 of Corydoras. These have a but curved block shaped head, most distinctively is a filament behind the barbel’s which can look like an additional barbel.
Aspidoras: Always known as Aspidoras but excludes Gastrodermis pauciradiatus.
Scleromystax: Sometimes known as bearded Corydoras, the only to show really clear sexual dimorphism through odontodes.
Gastrodermus: Originally the C. elegans group, lineage 5 and the microcorydoras such as G. pygmeaus and G. hastatus. Much shorter and rounder in the head but still slightly trinagular unlike Hoplisoma.
Osteogaster: Originally the C. aeneus group, contains the bronze, O. aeneus. Rounded head, short.
Brochis: This is a much larger group, likely for the time being. Contains the whole of lineage 8 from what was Brochis to species such as C. arcuatus. Similar to Corydoras the majority have large heads but these lack the curvature of Corydoras, similarly contains many large members.
Hoplisoma: Lineage 9, another very popular genus from the trade. Very rounded head but much shorter then Osteogaster.
Corydoradinae head shapes
The clearest way to see the difference between the different genera is their head shapes, there is quite the diversity as shown in the above figure. The majority of these genera can be identified from that head shape in my opinion. Brochis seems to display the widest range of morphological disparity in head shape but this can be expected from such a large genus.
Why is this revision important?
This revision reflects our current knowledge of the group Corydoradinae, both regarding morphological and molecular information. Rather then have one large category which doesn’t describe the morphological diversity or any phylogenetic information, these genera are easier to navigate particularly for a hobbyist. Not just does their morphology differ but likely their care. I already know Corydoras and some Brochis display territorial behavior which is not reflective of all groups, from a hobbyist view point knowing the genera helps avoid this, or avoid larger species.
Revisions are normal in science as technology and knowledge advances, they aren’t done for no reason or for publication. Over time we have developed techniques that have helped us understand the evolution of fishes better and therefore give them more accurate names that reflect this evolution. Technology such as Computerized tomography (CT) scanning, extracting DNA from formalin preserved/historical specimens, gene expression, developmental techniques etc. We can now understand fishes in a way we never could before, it means that change is inevitable but it’s the path to understanding evolution, biodiversity and reflecting that in how we name species, genera, families etc.
This work was done by the scientists; Angelica C. Dias, Luiz F. C. Tencatt, Fabio F. Roxo, Gabriel de Souza da Costa Silva, Sérgio A. Santos, Marcelo R. Britto, Dr. Martin I. Taylor and Dr. Claudio Oliveira.
No doubt our knowledge of Corydoradinae wouldn’t be the same without the efforts of Steven Grant (Catfishes of the World) and Ian Fuller who runs Corydoras World, both have done a lot for bridging the gap between the hobbyist and the scientist. Corydoras World I can certainly recommend for anyone who wants to learn more about this curious group of fishes.
What species are what?
To save time I wont cite authors, but this should ideally be done. I wont include C and CW numbers as it’ll make the lists much much longer.
Corydoras
Corydoras acutus
Corydoras amapaensis
Corydoras areio
Corydoras aurofrenatus
Corydoras blochi
Corydoras caramater
Corydoras cervinus
Corydoras coriatae
Corydoras cortesi
Corydoras desana
Corydoras filamentosus
Corydoras fowleri
Corydoras fulleri
Corydoras geoffroy
Corydoras maculifer
Corydoras narcissus
Corydoras negro
Corydoras ourastigma
Corydoras oxyrhynchus
Corydoras pastazensis
Corydoras saramaccensis
Corydoras sarareensis
Corydoras semiaquilus
Corydoras septentrionalis
Corydoras serratus
Corydoras simulatus
Corydoras solox
Corydoras spilurus
Corydoras stenocephalus
Corydoras treitlii
Corydoras vittatus
Corydoras zawadzkii
Aspidoras
Aspidoras albater
Aspidoras aldebaran
Aspidoras azaghal
Aspidoras belenos
Aspidoras brunneus
Aspidoras carvalhoi
Aspidoras depinnai
Aspidoras fuscoguttatus
Aspidoras gabrieli
Aspidoras kiriri
Aspidoras lakoi
Aspidoras maculosus
Aspidoras mephisto
Aspidoras poecilus
Aspidoras psammatides
Aspidoras raimundi
Aspidoras rochai
Aspidoras velites
Scleromystax
Scleromystax barbatus
Corydoras lacerdai
Scleromystax macropterus
Scleromystax prionotos
Scleromystax reisi
Scleromystax salmacis
Scleromystax virgulatus
Gastrodermus
Gastrodermus bilineatus
Gastrodermus elegans
Gastrodermus gracilis
Gastrodermus guapore
Gastrodermus hastatus
Gastrodermus latus
Gastrodermus mamore
Gastrodermus nanus
Gastrodermus napoensis
Gastrodermus nijsseni
Gastrodermus paucerna
Gastrodermus pauciradiatus
Gastrodermus pygmaeus
Gastrodermus undulatus
Osteogaster
Osteogaster aeneus
Osteogaster eques
Osteogaster hephaestus
Osteogaster maclurei
Osteogaster melanotaenia
Osteogaster rabauti
Osteogaster zygatus
Brochis
Brochis agassizii
Brochis amandajanea
Brochis ambiacus
Brochis approuaguensis
Brochis arcuatus
Brochis bethanae
Brochis bifasciatus
Brochis britskii
Brochis brittoi
Brochis condiscipulus
Brochis costai
Brochis crimmeni
Brochis crypticus
Brochis delphax
Brochis deweyeri
Brochis difluviatilis
Brochis ephippifer
Brochis garbei
Brochis geryi
Brochis gomezi
Brochis haraldschultzi
Brochis heteromorphus
Brochis imitator
Brochis incolicana
Brochis isbrueckeri
Brochis lamberti
Brochis leopardus
Brochis multiradiatus
Brochis noelkempffi
Brochis ornatus
Brochis orphnopterus
Brochis pantanalensis
Brochis pinheiroi
Brochis pulcher
Brochis reticulatus
Brochis robineae
Brochis robustus
Brochis seussi
Brochis sodalis
Brochis spectabilis
Brochis splendens
Brochis sychri
Brochis virginiae
Hoplisoma
Hoplisoma acrensis
Hoplisoma adolfoi
Hoplisoma albolineatum
Hoplisoma amphibelum
Hoplisoma apiaka
Hoplisoma araguaiaensis
Hoplisoma armatum
Hoplisoma atropersonatum
Hoplisoma axelrodi
Hoplisoma baderi
Hoplisoma benattii
Hoplisoma bicolor
Hoplisoma boehlkei
Hoplisoma boesemani
Hoplisoma bondi
Hoplisoma breei
Hoplisoma brevirostris
Hoplisoma burgessi
Hoplisoma carlae
Hoplisoma caudimaculatum
Hoplisoma cochui
Hoplisoma colossus
Hoplisoma concolor
Hoplisoma copei
Hoplisoma coppenamensis
Hoplisoma cruziensis
Hoplisoma davidsandsi
Hoplisoma diphyes
Hoplisoma duplicareum
Hoplisoma ehrhardti
Hoplisoma esperanzae
Hoplisoma evelynae
Hoplisoma eversi
Hoplisoma flaveolum
Hoplisoma froehlichi
Hoplisoma gladysae
Hoplisoma gossei
Hoplisoma granti
Hoplisoma griseum
Hoplisoma gryphus
Hoplisoma guianensis
Hoplisoma habrosum
Hoplisoma julii
Hoplisoma kanei
Hoplisoma knaacki
Hoplisoma lacrimostigmata
Hoplisoma leucomelas
Hoplisoma longipinnis
Hoplisoma loretoensis
Hoplisoma loxozonum
Hoplisoma lymnades
Hoplisoma melanistium
Hoplisoma melini
Hoplisoma metae
Hoplisoma micracanthus
Hoplisoma microcephalum
Hoplisoma multimaculatum
Hoplisoma nattereri
Hoplisoma oiapoquensis
Hoplisoma ortegai
Hoplisoma osteocarum
Hoplisoma paleatus
Hoplisoma panda
Hoplisoma paragua
Hoplisoma parallelum
Hoplisoma pavanelliae
Hoplisoma petracinii
Hoplisoma polystictum
Hoplisoma potaroensis
Hoplisoma punctatum
Hoplisoma revelatum
Hoplisoma reynoldsi
Hoplisoma sanchesi
Hoplisoma schwartzi
Hoplisoma similis
Hoplisoma sipaliwini
Hoplisoma steindachneri
Hoplisoma sterbai
Hoplisoma surinamensis
Hoplisoma trilineatum
Hoplisoma tukano
Hoplisoma urucu
Hoplisoma weitzmani
Hoplisoma xinguensis
References:
Alexandrou, M. A., Oliveira, C., Maillard, M., McGill, R. A., Newton, J., Creer, S., & Taylor, M. I. (2011). Competition and phylogeny determine community structure in Müllerian co-mimics. Nature, 469(7328), 84-88.
Britto, M. R. (2003). Phylogeny of the subfamily Corydoradinae Hoedeman, 1952 (Siluriformes: Callichthyidae), with a definition of its genera. Proceedings of the Academy of Natural Sciences of Philadelphia, 153(1), 119-154.
Dias, A. C., Tencatt, L. F., Roxo, F. F., Silva, G. D. S. D. C., Santos, S. A., Britto, M. R., … & Oliveira, C. (2024). Phylogenomic analyses in the complex Neotropical subfamily Corydoradinae (Siluriformes: Callichthyidae) with a new classification based on morphological and molecular data. Zoological Journal of the Linnean Society, zlae053.
Shimabukuro-Dias, C. K., Oliveira, C., Reis, R. E., & Foresti, F. (2004). Molecular phylogeny of the armored catfish family Callichthyidae (Ostariophysi, Siluriformes). Molecular Phylogenetics and Evolution, 32(1), 152-163.
Little seems known about the diet of Corydoradinae, it might be due to a generalization of the genus but also a lack of understanding of their ecology. For scientists it is a relatively small genus from a small family, Callichthyidae which also contains genera such as Megalechis, Hoplosternum and Aspidoras.
Brochis bethanae CW006 also known as the Narcisso Corydoras taken at Maidenhead Aquatics, Ascot
There is no doubt that Corydoras feeds largely as a carnivore but as a term that is very vague. These fishes are hardly hunting down capybara that enter the water or swarming round the carcasses of fishes. Carnivory purely just refers to the fact an animal eats an animal, and what is defined as an animal is just Animalia which is a gigantic category of organisms. Animalia covers from the simple sponges and corals all the way to molluscs, mammals and fishes, it includes some strange organisms as well like bryozoa and jellyfish. Each of these animals will have different nutritional compositions, some toxins but also accessibility. It is well known that insectivores (carnivores that specialize on insects) are not able to access nutrition from fishes, mammals etc. efficiently (Žák et al., 2022), this also increases nitrogenous waste. There might be other aspects of nutrition commonly missed such as perhaps the importance of chitin? It’s very difficult to digest, too much and the food item wont be processed, too little might result in blockages. It reminds me very much of the bloodworm used in the aquarium trade having a strong chitin casing compared to the chromatid larvae the fishes are likely eating in the wild. I have seen bloodworm pass out the fish as if it hadn’t been eaten at all, even carnivorous fishes. Not just does carnivory cover a wide range of different nutritional profiles and species but also modes of feeding. It is very different to catch and feed on a whole fish as it is to maybe feed on scales (lepidophagy), break down snails (durophagy), maybe extract a snail from it’s shell etc. Carnivory is so obviously diverse compared to herbivory but it still is best visualized like a field of grass, all grass specialists but put sheep, cattle and horses out and they will all feed on very different parts of the grass.
What I am emphasizing is not to generalize any dietary category and just fall to the general diets for fishes. Carnivory is just a man made category regardless and doesn’t reflect realistically aquatic dietary niches.
Corydoradinae has previously been identified as an omnivore by Nijssen (1970) although only used aquarium fishes as evidence. This paper records them feeding on fallen leaves of which I find particularly strange. Although a particular fondness for invertebrates is noted, particularly tubifex and daphina, identifying the worms using their highly evolved sense of smell/taste. This record is later referenced in Alexandrou et al. (2011) but also noted that they feed on algae, insects, zooplankton and annelids. Algae I would not be surprised that is consumed but I do not believe they are targeting it, much like invertebrates are found in small numbers the guts of grazing Loricariids.
Isotope analysis was used to compare different lineages of Corydoras and identify any partitioning in where and what they feed on. Different lineages display divergent nutritional profiles between different genera of Corydoradinae based on head shape, eye placement and body depth. There is a clear difference between the diet of the longer snouted and shorter snouted Hoplisoma and Gastrodermus, I can assume due to the depths of substrate that can be exploited by either. These longer snouted, Corydoras are referenced as feeding on a lower trophic level (Alexandrou et al., 2011). This could infer on feeding on more algaes but maybe those lower trophic level invertebrates such as worms who would be lower down in the substrate then predatory invertebrates.
The shorter to medium snouted Hoplisoma paleatum, the peppered ‘Corydoras‘ is recorded as feeding largely on fly larvae such as chromatid’s with a small addition of nematode’s. Algae and plant fragments are recorded in the gut but near minimal volumes, less then substrate ingested (Bertora et al., 2021).
So the picture of what Corydoradinae eat in the wild is unclear.
The Dietary Morphology of Corydoras
Figure 1: Hoplisoma trilineatum skeletal anatomy produced by computerized tomography (CT) scanning. Produced by: Lowe, A., Summers, A. P., Walter, R. P., Walker, S., & Paig-Tran, E. M. (2021). Scale performance and composition in a small Amazonian armored catfish, Corydoras trilineatus. Acta Biomaterialia, 121, 359-370.
Again, an unclear topic. We know Corydoradinae have oral jaws (Fig 1) at the front of the head humongous with our the general vertebrate jaw. These oral jaws contain teeth (Huysentruyt et al., 2011) but there seems to have been no exploration of the diversity of these teeth. These oral jaws are very similar to other invertivores (feeds on invertebrates), being elongate to extract food items out of crevices or the substrate.
What is not researched is the secondary pair of jaws found in most fishes, the pharyngeal jaws. While the oral jaws in fishes are often involved in prey capture, the pharyngeal jaws are involved in prey processing, so the grinding and breaking down. These are at the back of the mouth so aren’t obvious but when you see a fishes head move after feeding it’s likely those jaws are moving. Corydoradinae do have pharyngeal jaws, they contain teeth (Huysentruyt et al., 2011) but we have no idea how this morphology differs across Corydoradinae. In Osteogaster aenea Huysentruyt et al. (2011) identified elongate pharyngeal teeth which would confirm that at least O. aeneus is not evolved to feed on snails but the jaws do seem some what robust. It contrasts from those species that feed on algae to any extent who seem to have much more simplistic and often bladed pharyngeal anatomy. Most research into pharyngeal anatomy focuses on cichlids of which might not be the best reference given differential feeding behaviour.
Head shape can tell a lot about the fish, these fishes have such inferiorly facing mouths will be feeding around the substrate. What is more interesting is the shape of the snout mentioned earlier but we know so little about it. Those elongate snouts certainly allow the fish to dig deeper for food but how it effects them we only have clues.
What should I feed my Corydoras?
These are certainly not feeding on fishes, there is also the misconception that because fishes die that all fishes have access to them. Generally weaker individuals would be picked up by predators before they die and any dead fish would be more quickly exploited by species evolved to detect and quickly feed on carcasses. So fish meal is logically best avoided. These fishes also do not feed on plants so cereals are certainly of little use. This means those general diets are not great for them, most containing fish meal, cereals, vegetables etc. usually in that order.
Luckily for carnivores there is a few options for insect based foods. Fish Science is generally good and they offer some diversity. Fluval bug bites can be okay but still has quite a lot of fish meal and cereals in it. Repashy bottom scratcher is certainly worth looking at, being a gel diet you can add additional ingredients to it. I wouldn’t be afraid to use either Fish Science or Repashy bottom scratcher as the basis of the diet then frozen and freeze dried foods to build on it creating a more well rounded diet. Certainly live foods are worth looking at and would offer a lot of enrichment.
Corydoradinae are very forgiving regarding diets so experimenting is certainly possible.
Frozen foods are great but often will not contain all of the nutrition a species might require. In the wild most fishes will feed on hundreds of species and the narrow range of frozen foods available likely doesn’t compare nutritionally. These are great as enrichment or an addition to a fishes diet but not as a complete diet.
Protein Blisters
This is something that will always need mentioning regarding diets. I am not convinced it is caused by too much protein, even protein that isn’t absorbed/taken up by the fish e.g. excreted as nitrogenous waste. There is no evidence either way, it is clear bacteria can cause blisters and cysts but it can’t be said every cyst is caused by them without exploring further. Some frozen/live foods such as bloodworm can harbor Aeromonas (Senderovich et al., 2008) so it is difficult to make assumptions but there is little research on that.
A suggestion has been a form of gas bubble disease, true gas bubble disease as I call it though is caused from supersaturation of gases in the water, or some change in pressure resulting in bubbles forming in tissues very rapidly like the bends. It kills extremely rapidly as those bubbles form and burst blood vessels. There are other similar diseases like I have seen extreme algae growth associated, not proven though, to cause bubbles in some fishes but doesn’t kill and they are very localized.
We see similar in Loricariids, also armored and personally I would associate it with an infection of some kind. Blisters are tricky though as it’s such a general pathological symptom it could mean anything.
References:
Alexandrou, M. A., Oliveira, C., Maillard, M., McGill, R. A., Newton, J., Creer, S., & Taylor, M. I. (2011). Competition and phylogeny determine community structure in Müllerian co-mimics. Nature, 469(7328), 84-88.
Bertora, A., Fontanarrosa, M. S., Grosman, F., Sanzano, P., & Rosso, J. J. (2021). Trophic ecology of the Neotropical tolerant fish Corydoras paleatus under the influence of contrasting environmental conditions in a prairie stream. Anais da Academia Brasileira de Ciências, 93, e20200981.
Huysentruyt, F., Geerinckx, T., Brunain, M., & Adriaens, D. (2011). Development of the osteocranium in Corydoras aeneus (Gill, 1858) Callichthyidae, Siluriformes. Journal of Morphology, 272(5), 573-582.
Lowe, A., Summers, A. P., Walter, R. P., Walker, S., & Paig-Tran, E. M. (2021). Scale performance and composition in a small Amazonian armored catfish, Corydoras trilineatus. Acta Biomaterialia, 121, 359-370.
Nijssen, H. (1970). Revision of the Surinam catfishes of the genus Corydoras Lacépède, 1803 (Pisces, Siluriformes, Callichthyidae). Beaufortia, 18(230), 1-75.
Senderovich, Y., Gershtein, Y., Halewa, E., & Halpern, M. (2008). Vibrio cholerae and Aeromonas: do they share a mutual host?. The ISME journal, 2(3), 276-283.
Žák, J., Roy, K., Dyková, I., Mráz, J., & Reichard, M. (2022). Starter feed for carnivorous species as a practical replacement of bloodworms for a vertebrate model organism in ageing, the turquoise killifish Nothobranchius furzeri. Journal of Fish Biology, 100(4), 894-908.