Many fishkeepers are after something unique and usual for their aquarium and sadly many of what we call oddballs are usually quite large. There are always exceptions to this and these only need to be encouraged.
The eel issue
Before exploring Macrognathus circumcinctus I should explain the issue with the word eel. Eel is a common name that realistically describes anything anguilliform in shape. Anguilliform is a fish shape which refers to that elongate body shape and would generally use that body for locomotion. This is easily confused with Anguilliformes, true eels are quite distantly related from what we see sold as freshwater eels with the exception of the large Gymnothorax polyuranodon (Lifestyle is debated; Ebner et al., 2011; Ebner et al., 2019) and G. tile (actually brackish). Eels are found in many clades from the eel loaches in Pangio and Vaillentella; to the eel catfishes in Channallabes. Gymnotiformes, true knifefishes are also described as either eels or having anguilliform morphology.
No group is referred to as eels as much as Synbranchiformes and this is where Macrognathus circumcintus places. Other popular ‘eel’ fishes are also found in this clade, Mastacembelus erythrotaenia (Fire eel), Mastacembelus armatus (tyre track eel) and if more interested in Asian cooking then Monopterus spp.(swamp eels). The South American swamp eel, Synbranchus spp. is particularly interesting given along with being obligate air breathers they are sequential/diandric hermaphrodites (Barros et al., 2017).
The Half Banded Spiny Eel, Macrognathus circumcinctus
Taxonomy: Previously Mastacembelus before being revised in 1984.
Locality: Widespread throughout South East Asia. GBIF (Global Biodiversity Information Facility) records this species largely in Thailand and Cambodia with a few reaching out into Vietnam, Malaysia, Borneo and Sumatra. Whether these represent the same species I don’t know.
Size: 9-13cm SL (Maharsi et al., 2024; Parenti & Lim, 2005), there is some report of a 28.5cm individual from Fishbase but this seems exceptional.
Habitat: It seems they prefer rocky habitats with reasonable currents (Maharsi et al., 2024), they prefer it shaded and it also seems to be largely turbid not by tannins but by sediment. These habitats probably have a lot of invertebrates feeding on the diversity of bacteria.
Temperature: 26.7-29.2 (Maharsi et al., 2024)
pH: 6.4-8.33 (Maharsi et al., 2024).
Diet: Invertebrates (Maharsi et al., 2024), maybe small fishes but I have seen mine actively feed on mushrooms. They can be quite quick ambush predators while also capable of searching for food amongst crevices. This invertebrate diet is pretty consistent throughout Macrognathus (Sarmah & Kardong, 2024). Ideally as a fishkeeper it would be worth aiming for a diet with an insect/invertebrate based dry/gel diet such as Repashy Bottom Scratcher Or Fluval Bug Bites. Building on that diet with a diversity of frozen and freeze dried foods. You can also try live foods such as earthworms on larger individuals but tubifex, blackworms, daphina etc. on smaller.
Hardscape: A lot of decor so they have places to hide and explore, a confident individual will come out but also if in a group it gives them space to hide should be required.
Substrate: While I’ve never experienced this species burying particularly and even less with age many will bury in the sand and it is therefore beneficial as enrichment to provide that for them.
Sociality, a problem?
While almost all websites on the species will record this species as social, it doesn’t take much to find out the picture is a little more complicated. In reality some are and it’s worth buying more if you can because if yours are social they will benefit from that interaction as basic enrichment. Mine and many others have M. circumcinctus who wont tolerate others of their species and even other eel shaped fishes, while I thought it was because even before I took on mine it was housed alone for potentially many years. Over time it’s become clear mine is not alone and they can even be aggressive from a younger age. They are very much individuals so it is likely that more then anything else.
References:
Barros, N. H. C., De Souza, A. A., Peebles, E. B., & Chellappa, S. (2017). Dynamics of sex reversal in the marbled swamp eel (Synbranchus marmoratus Bloch, 1795), a diandric hermaphrodite from Marechal Dutra Reservoir, northeastern Brazil. Journal of Applied Ichthyology, 33(3), 443-449.
Ebner, B. C., Donaldson, J. A., Courtney, R., Fitzpatrick, R., Starrs, D., Fletcher, C. S., & Seymour, J. (2019). Averting danger under the bridge: video confirms that adult small-toothed morays tolerate salinity before and during tidal influx. Pacific Conservation Biology, 26(2), 182-189.
Ebner, B. C., Kroll, B., Godfrey, P., Thuesen, P. A., Vallance, T., Pusey, B., … & Perna, C. N. (2011). Is the elusive Gymnothorax polyuranodon really a freshwater moray?. Journal of Fish Biology, 79(1), 70-79.
Maharsi, G. J., Waluyo, W., & Armando, E. (2024). Study of Ecobiological Characteristics of Spiny Eel (Mastacembelidae) in Elo River, Magelang, Central Java. JFMR (Journal of Fisheries and Marine Research), 8(3), 8-15.
Parenti, L. R., & Lim, K. K. (2005). Fishes of the Rajang Basin, Sarawak, Malaysia. The Raffles Bulletin of Zoology, 13, 175-208.
Sarmah, P., & Kardong, D. (2024). Biology Of Macrognathus Sp. With Special Reference To Macrognathus Pancalus (Hamilton) And Macrognathus Aral (Bloch And Schneider). Journal of Advanced Zoology, 45(1).
It is quite common for nitrates to be labelled as non-toxic to fishes and it is kind of accepted that nitrates have a lower toxicity to ammonia and nitrites, or more that these have a higher acute (short term) toxicity. Every aquarium community will give you a different maximum level of nitrate toxicity, this kind of reflects that nitrate toxicity is very much misunderstood and rarely fact checked. Not just is this topic rarely fact checked but often it is a lot more complex as it takes understanding the limitations of the scientific literature.
One of the beautiful angelfish, Pterophyllum scalare at the Keystone Clash 2024.
It is noted in the scientific literature that nitrate exposure is neglected (Monsees et al., 2017), so why do we focus on the lack of studies in the assumption it’s non-toxic with little science to say it is.
At the end of the day, we know nitrates is toxic. We don’t know the safe levels and therefore it is more ethical to assume that as little as possible is safer then more because we know that nitrates have no benefits. It is better to assume that negative for the sake of a little bit of hard work if it could benefit the animal.
There are definitely flaws in the idea nitrates are not toxic particularly when saying up to 20, 40, 100ppm are fine. Let me discuss each limitation to these claims:
Most studies focus on species that we do not keep, hardy species such as tilapia (Oreochromis; Monsees et al., 2017), carp, Salmonids (Yu et al., 2021) or Danio rerio. Many of these do not apply either due to temperature or just not closely related to anything we are discussing.
These studies are short term so look at acute exposure not long term chronic exposure (Monsees et al., 2017; Yu et al., 2021). If the studies only look at 30 days it cannot be assumed that the effect is the same at 5, 10 or 20 years.
Studies use distinct gradings of exposure, often wide values so it is not possible to understand effects between those values (Gomez Isaza & Cramp, 2020; 2021; Yu et al., 2021). If they only test from 10ppm onwards the effect below 10ppm is unknown (Cano-Rocabayera et al., 2019) and if effects are only noted at 50ppm but they don’t test between that and 10ppm it cannot be assumed that 50ppm is a minimum level.
No study looks at all effects of exposure, particularly physiological effects so could miss other effects of the treatments exposed (Cano-Rocabayera, 2019; Monsees et al., 2017; Yu et al., 2021).
Many people focus on lethal effects in the short term and lethal doesn’t mean it isn’t causing other damage.
Why these limitations? Well the funding understandably is highest for food fishes and these do not need to be kept long term particularly on the scale for fishkeeping, along with that the aim is often not reproduction.
Lasiancistrus sp. obtained from Acres Aquatics, Warminster UK.
We know that nitrate is a pollutant that requires long term, chronic exposure (Yu et al., 2021) so this is where the limitation in understanding arises. When it comes to exposure levels 0ppm is usually the control to the next exposure level is tested there are effects on the fishes physiology (Kellock et al., 2018; Gomez Isaza & Cramp, 2021) that many aquarists would not be able to test for. While not always statistical there seems to always be effect but not lethal till very very high concentrations e.g. 600ppm, Regardless it does effect adaptability to change (Presa et al., 2022).
It does seem in every study there is a difference between low level exposure and no exposure in the short term in those cited but it is not significant, what about long term? If we think this is short term exposure causing some damage but over time and years, is recovery possible? I’d assume likely not.
There is never really any support for any one value to be the toxic level for many or any fishes (Isaza et al., 2020; Presa et al., 2011; Villa-Villaseñor et al., 2022; in general any of the citations).
What are the effects of nitrates?
Natural habitats are known as oligotrophic and normally have very little nitrates that is close to the undetectable level without a pollution event. Clearly captive fishes have a little more adaptability but many of the fishes we keep are wild caught and does it depend on the generation (Isaza et al., 2020).
Male red cover discus, Symphysodon sp. x. with Xiphophorus maculatus x. X. variatus
Test kit reliability.
The crutch a reliance on nitrates being low levels before water changes relies on reliable test kits. Dip stick, paper test kits have always been argued as unreliable, giving both false negatives and false positives although when first opened many will note they do show some reliability. Liquid test kits are a little bit more complicated, obviously we can’t calibrate them personally as we have nothing to calibrate them against as a hobby and similarly if using a digital system.
We think our liquid test kits are 100% reliable but they do expire and once opened it’s tricky to say when this happens. I generally say liquid test kits should be replaced annually as there can be false results after that depending on brand and product. But I have seen some test kits newly opened, long before their expiry date give both false negatives and positives for nitrates. So, why rely on these for water changes?
Conclusions
I think this argument is similar to UVB to produce vitamin D in reptiles, there has always been the idea that UVB is not needed for certain reptiles, they are nocturnal or consume whole mammals who provide vitamin D. Yet over time there has been evidence that even in these animals there is a benefit to providing Vitamin D. As this is not a reptile website and to save all the citations I will forward the Reptile Lighting Facebook group: https://www.facebook.com/groups/384134861721116 This group provides many citations.
Water changes are vital but also better to be cautious when it comes to test kits and go on the precaution of maybe more nitrates then you actually have. It’s also worth testing your tap water.
While we don’t know the long term damage of low level nitrates it’s worth acting on the precaution.
References:
Cano-Rocabayera, O., De Sostoa, A., Padrós, F., Cardenas, L., & Maceda-Veiga, A. (2019). Ecologically relevant biomarkers reveal that chronic effects of nitrate depend on sex and life stage in the invasive fish Gambusia holbrooki. Plos one, 14(1), e0211389.
Gomez Isaza, D. F., Cramp, R. L., & Franklin, C. E. (2020). Simultaneous exposure to nitrate and low pH reduces the blood oxygen-carrying capacity and functional performance of a freshwater fish. Conservation physiology, 8(1), coz092.
Gomez Isaza, D. F., Cramp, R. L., & Franklin, C. E. (2021). Exposure to nitrate increases susceptibility to hypoxia in fish. Physiological and Biochemical Zoology, 94(2), 124-142.
Isaza, D. F. G., Cramp, R. L., & Franklin, C. E. (2020). Living in polluted waters: A meta-analysis of the effects of nitrate and interactions with other environmental stressors on freshwater taxa. Environmental Pollution, 261, 114091.
Kellock, K. A., Moore, A. P., & Bringolf, R. B. (2018). Chronic nitrate exposure alters reproductive physiology in fathead minnows. Environmental Pollution, 232, 322-328.
Monsees, H., Klatt, L., Kloas, W., & Wuertz, S. (2017). Chronic exposure to nitrate significantly reduces growth and affects the health status of juvenile Nile tilapia (Oreochromis niloticus L.) in recirculating aquaculture systems. Aquaculture Research, 48(7), 3482-3492.
Presa, L. S., Neves, G. C., Maltez, L. C., Sampaio, L. A., Monserrat, J. M., Copatti, C. E., & Garcia, L. (2022). Acute and sub‐lethal effects of nitrate on haematological and oxidative stress parameters of juvenile mullet (Mugil liza) in freshwater. Aquaculture Research, 53(9), 3346-3357.
Villa-Villaseñor, I. M., Yáñez-Rivera, B., Rueda-Jasso, R. A., Herrera-Vargas, M. A., Hernández-Morales, R., Meléndez-Herrera, E., & Domínguez-Domínguez, O. (2022). Differential sensitivity of offspring from four species of goodeine freshwater fish to acute exposure to nitrates. Frontiers in Ecology and Evolution, 10, 1014814.
Yu, J., Xiao, Y., Wang, Y., Xu, S., Zhou, L., Li, J., & Li, X. (2021). Chronic nitrate exposure cause alteration of blood physiological parameters, redox status and apoptosis of juvenile turbot (Scophthalmus maximus). Environmental Pollution, 283, 117103.
This is a big question when it comes to keeping Loricariids in the aquarium as more then often people want to spawn the fishes but sometimes people just want to name them.
What is the sex determination methods in Loricariidae?
This is actually a more vast discussion then some other groups like mammals. In fact Loricariid’s use a range of karyotypes to determine sex from the more well known ZW and XY to multiple sex chromosomes. Ancistrus (Bristlenoses) is a large genus representing over 60 spp. (Eschmeyer, 2025), this genus does show multiple sex determination methods and some not as well known such as ZZ/ZW1W2 in Ancistrus clementinae (Nirchio et al., 2023) or ZZ/ZW in Ancistrus ranunculus (Oliveira et al., 2007). It seems many genera show a lot of diversity (Sassi et al., 2023) and this is possibly a large barrier to hybridization and maybe could lead to speciation.
What do you need to be able to identify the sex of your Loricariid?
A mature fish, unless the fish has reached sexual maturity it likely wont show many sexually dimorphic features although there are slight exceptions at the smaller level or during dissection. Largely this will mean close to fully grown and/or clearer odontode growth. Some species this might take a year such as some of the more common Ancistrus or 5+ years like the Hemiancistrus medians group (e.g. Panaque, Baryancistrus etc.)
A bowl, container or polybox can be useful to examine around the fish particularly below. Fill this with the tank water, do not do it during acclimation. It can be done in store.
The genital papilla and the urogenital pore are the same thing, this is a combined organ where the fish passes waste but also the gametes (eggs and sperm). This is not the same for all Loricariids where the genital pore is separate from the anus such as in Neoplecostomus. It is clearly on the abdomen, shape is normally ambiguous in juveniles.
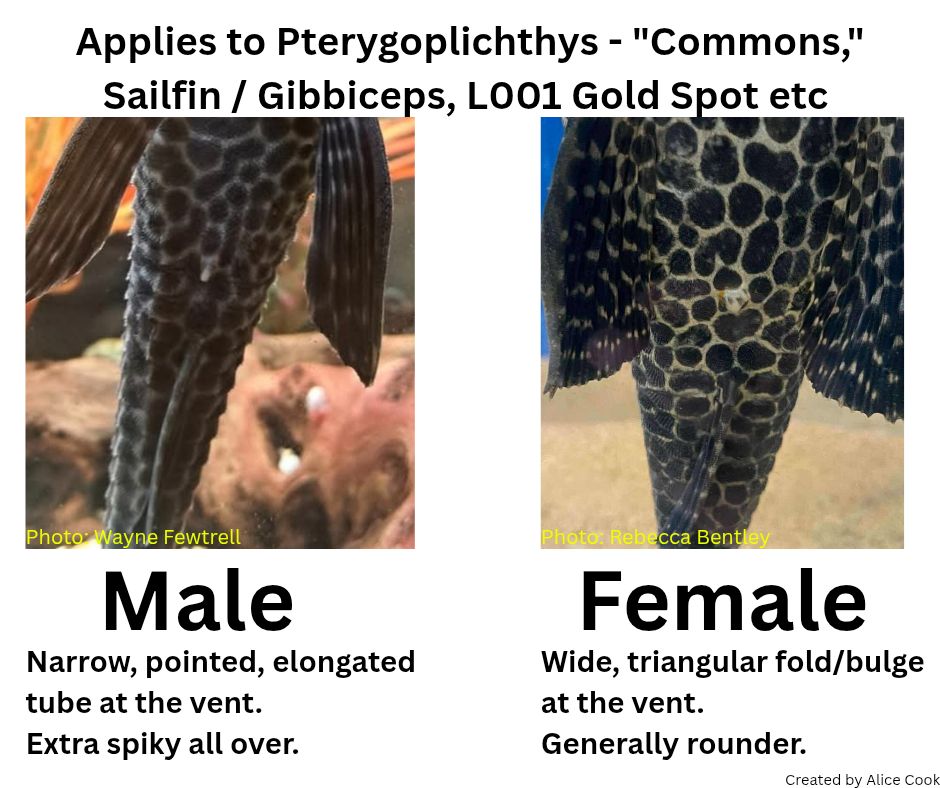
The shape of the genital papilla is the most reliable method to sex Lorcariids, it’s most easily stated as V shaped in males and U in females. This can be tricky to see in some genera which have more elongate genital papilla such as the Pterygoplichthys in figure 2.
Figure 2: created by In the Bag Tropical Fish UK, Alice Cook. Depicting Pterygoplichthys pardalis and Pterygoplichthys gibbiceps.Figure 4: Mature female Baryancistrus chrysolomus, not showing the clearest maturity.
Some other genera the females are much wider so it is much more square as above. The best method is to see multiple individuals and compare between them which can be done against the glass and taking photos or even within a container to take a more up close look.
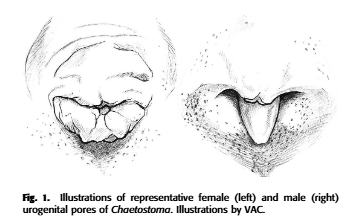
Figure 5: The genital papilla of Chaetostoma as explained by Lujan et al. (2015) in Lujan, N. K., Meza-Vargas, V., Astudillo-Clavijo, V., Barriga-Salazar, R., & López-Fernández, H. (2015). A multilocus molecular phylogeny for Chaetostoma clade genera and species with a review of Chaetostoma (Siluriformes: Loricariidae) from the Central Andes. Copeia, 103(3), 664-701.
I find this method the most reliable particularly as there is less individual variation between individuals and also depends less on the age of the fish. The shape can be applied also to Loricariinae (whiptail catfishes) and Hypoptopominae (Otocinclus, Parotocinclus, Hypoptopoma etc.).
The final part of the genital papilla when it comes to sexing Loricariids is spotting around this region in females. These papilla/spots can be green or yellow in colour and can range from one to many in number, once visible to the eye it hints maturity in the females. While it is very obvious in Hypostominae (traditional pleco’s) it is unclear as to if these are present in the other subfamilies even the distantly related but similarly looking Pseudancistrus genisetiger (slate pleco’s) and Rhinelepinae. Although I have been told by Fauna Tropica (https://www.faunatropica.eu/) that these spots can be seen under a microscope and maybe a macrolens before maturity even as younger juveniles.
Figure 6: Genital papilla of a female Baryancistrus chrysolomus.
Body shape
I am not a massive fan of using body shape as it can depend on many factors. Using how plump the fish is does depend on how well the fish has been fed but also females once the female has released eggs can suddenly reduce weight.
Head shape does seem the most reliable method regarding the general anatomy but can be limited when it comes to stunted fish but there is also a lot of individual variation. In general it is assumed females display more elongate heads whereas males are shorter and wider, this is a trend we do see in other fishes.
Figure 7: Body shape in two different Scobinancistrus species but clearly shows the sexual dimorphism.
Personally when it comes to sexing individuals from photos I am less of a fan of this method as it does rely even more on angle of the photo but also maturity. You can see a larger difference in some genera then others and some species you might not see it at all.
Odontodes
Figure 8: Opercular odontodes on Ancistrus ranunculus.
Odontodes are the external teeth that cover Loricariids, this does include everything from Otocinclus to Farlowella to Hypancistrus. In some species they can be enlarged in certain areas and this is known as hypertrophied, further in some these odontodes can be sexually dimorphic. Odontodes as a secondary sexual characteristic though are not a rule and there are many exceptions, in addition they can be seasonal so males might have reduced ones seasonally and females can have larger ones, it depends on the genus.
Odontodes are most useful in the Peckoltia clade, this includes Hypancistrus, Peckoltia, Panaqolus, Pseudoqolus, Ancistomus and Scobinancistrus. Although Scobinancistrus and species such as Peckoltia sabaji do not have particularly sexually dimorphic odontodes and I recommend using the genital papilla. These odontodes are hypertrophied in all individuals at the gill opercular although can be larger in males. The most obvious method with this clade is hypertrophied odontodes on the caudal peduncle and pectoral fin of the males.
Figure 10: Baryancistrus demantoides (high finned green phantom), female.
When using this trait be careful with other clades, some such as the Hemiancistrus medians group which includes Hemiancistrus medians, some other Hemiancistrus (this genus requires revision), Panaque, Parancistrus, Baryancistrus and some Spectracanthicus (again another genus that requires revision). In these clades I find females can grow large odontodes seasonally and this was very evident to me in a clearly female Baryancistrus demantoides (Fig 10). The key thing to note about odontodes is they can be shed.
Figure 11: Farlowella vittata group pictured at Aqualife, Leyland.
Another clade that easily gets forgotten is Loricariinae, while genital papilla are a clear way to sex many genera within this subfamily. Odontodes can provide an additional quick way to sex many particularly Farlowellini (Farlowella, Sturisoma and Sturisomatichthys mostly; Fig 11). These are reliably hypertrophied around the head and/or rostrum in this clade.
Tentacles
This is an exclusive trait to the genus Ancistrus and the species Lasiancistrus tentaculatus. These are fleshy growths derived from the odontode sheaths (Sabaj et al., 1999), which might explain why sexually dimorphism is shown as related clades such as Lasiancistrus shows some clearer dimorphism in the odontodes. In Ancistrus while in the common bristlenose the males display larger tentacles whereas the females display little to none, there are Ancistrus where females have large tentacles, some which have none and some where the size is the same. Not to be confused with odontodes which is why it is best not to refer to them as spines or bristles.
Some factors that are often myths when it comes to sexing Loricariids
Behaviour
It is a common misconception that males are more aggressive then females and often this has no grounding in experience or science, it’s often an assumption. Males and females do have different territories as males are involved in the brood care whereas females roam but where species are territorial it is in both sexes. In territorial species aggression is shown between and within the sexes. Often this idea of males being more aggressive is based in the coy female myth (Milam, 2013; Rosvall, 2013), many scientists have disproved this but Lucy Cooke makes some good approachable books to the topic. Females in Loricariids have no reason to be less aggressive. Personal experience I’ve seen aggression from both and if anything where females tend to roam their aggression is wider spread then close to the caves where males dwell.
The other myth is that females will not use caves, particularly as juveniles they will definitely hide a lot but even as adults females use caves as refuges, they might even have a preferred cave. Males use the caves to spawn in Hypostominae and some Loricariinae but not all do and those that like crevices will use them.
Conclusion
It can be really tricky to identify whether your Loricariid is male or female. Hopefully this offers some ideas to help sex your fishes. I tend to recommend using the abdomen of the fish as I feel this is most reliable and doesn’t leave space for any amounts of individual variation.
Lujan, N. K., Meza-Vargas, V., Astudillo-Clavijo, V., Barriga-Salazar, R., & López-Fernández, H. (2015). A multilocus molecular phylogeny for Chaetostoma clade genera and species with a review of Chaetostoma (Siluriformes: Loricariidae) from the Central Andes. Copeia, 103(3), 664-701.
Milam, E. L. (2013). Making males aggressive and females coy: Gender across the animal-human boundary. In Women, Science, and Technology (pp. 206-222). Routledge.
Nirchio, M., Oliveira, C., de Bello Cioffi, M., de Menezes Cavalcante Sassi, F., Valdiviezo, J., Paim, F. G., … & Rossi, A. R. (2023). Occurrence of sex chromosomes in fish of the genus Ancistrus with a new description of multiple sex chromosomes in the Ecuadorian endemic Ancistrus clementinae (Loricariidae). Genes, 14(2), 306.
Oliveira, R. R. D., Feldberg, E., Anjos, M. B. D., & Zuanon, J. (2007). Karyotype characterization and ZZ/ZW sex chromosome heteromorphism in two species of the catfish genus Ancistrus Kner, 1854 (Siluriformes: Loricariidae) from the Amazon basin. Neotropical Ichthyology, 5, 301-306.
Rosvall, K. A. (2013). Proximate perspectives on the evolution of female aggression: good for the gander, good for the goose?. Philosophical Transactions of the Royal Society B: Biological Sciences, 368(1631), 20130083.
Sabaj, M. H., Armbruster, J. W., & Page, L. M. (1999). Spawning in Ancistrus (Siluriformes: Loricariidae) with comments on the evolution of snout tentacles as a novel reproductive strategy: larval mimicry. Ichthyological Exploration of Freshwaters, 10(3), 217-229.
Sassi, F. D. M. C., Deon, G. A., Sember, A., Liehr, T., Oyakawa, O. T., Moreira Filho, O., … & Cioffi, M. D. B. (2023). Turnover of multiple sex chromosomes in Harttia catfish (Siluriformes, Loricariidae): a glimpse from whole chromosome painting. Frontiers in Genetics, 14, 1226222.
The use of various items produced by bees is popular within our daily lives, the most commonly used product being honey. The health benefit of products related to bees have been noticed for many hundreds maybe thousands of years from anti-microbial properties to general health. Honey maybe isn’t the most practical in the aquarium but two products you might find; bee pollen and propolis.
What is bee pollen?
Bee pollen is literally just the pollen collected by the bees. It is generally collected by forcing the bees to enter the hive through smaller then normal holes so the pollen drops below to be collected. This possibly puts a lot of pressure on the hive depending on how this is done.
What is the purpose of bee pollen in the aquarium?
Bee pollen is a dietary additive, there is a rising popularity in regards of this ingredient for people and perhaps this is the source of the idea. While it is an interesting ingredient and for those that might feed on fruit there could be a benefit. The actual benefits in general are debatable, with an omnivorous/carnivorous Clarias sp. catfish there were many physiological benefits shown up to 1% addition (Nowosad et al., 2022). When bee pollen was added to the diet of the insectivore zebrafish, Danio rerio there is no shown benefits to physiology but improve viral resistance (Di Chiacchio et al., 2021). In Nile tilapia, Oreochromis niloticus liver regeneration is shown to occur when fed bee pollen (Picoli et al., 2019).
What is propolis?
Propolis is the compounds and structures that seal the hive and the waxy structures that the hive is built on. It originates from the waxy areas of plants such as buds.
What is the purpose of propolis in the aquarium?
It’s not particularly common in aquariums but you might find the produce available more within the koi hobby. This product is used largely to seal wounds not just for protection against microbes entering them but also if you want to seal in a treatment. There does seem to be little research into the topic although some of the compounds do show promise when it comes to wound healing (Wibowo et al., 2021) and nutrition (Farag et al., 2021). More importantly there seems to be no papers onto any toxicity of propolis which is somewhat promising but could also be useful to cross out.
Conclusion
A very brief article I guess as to what could apply to the aquarist, it seems there needs a lot more research but I wouldn’t be afraid to use it. Personally I think bee pollen could be great for promoting feeding in fishes due to the sugars present. It’s something I think we could explore further.
References:
Di Chiacchio, I. M., Paiva, I. M., de Abreu, D. J., Carvalho, E. E., Martínez, P. J., Carvalho, S. M., … & Murgas, L. D. S. (2021). Bee pollen as a dietary supplement for fish: Effect on the reproductive performance of zebrafish and the immunological response of their offspring. Fish & Shellfish Immunology, 119, 300-307.
Farag, M. R., Abdelnour, S. A., Patra, A. K., Dhama, K., Dawood, M. A., Elnesr, S. S., & Alagawany, M. (2021). Propolis: Properties and composition, health benefits and applications in fish nutrition. Fish & Shellfish Immunology, 115, 179-188.
Nowosad, J., Jasiński, S., Arciuch-Rutkowska, M., Abdel-Latif, H. M., Wróbel, M., Mikiewicz, M., … & Kucharczyk, D. (2022). Effects of bee pollen on growth performance, intestinal microbiota and histomorphometry in African catfish. Animals, 13(1), 132.
Picoli, F., Lopes, D. L. D. A., Zampar, A., Serafini, S., Freccia, A., Veronezi, L. O., … & Emerenciano, M. G. C. (2019). Dietary bee pollen affects hepatic–intestinal histomorphometry of Nile tilapia fingerlings. Aquaculture Research, 50(11), 3295-3304.
Wibowo, I., Utami, N., Anggraeni, T., Barlian, A., Putra, R. E., Indriani, A. D., … & Ekawardhani, S. (2021). Propolis can improve caudal fin regeneration in zebrafish (Danio rerio) induced by the combined administration of Alloxan and glucose. Zebrafish, 18(4), 274-281.
Eels constantly cause fascination within aquarists but many true eels, Anguilliformes are simply too large for the majority of aquarists. A much smaller but fascinating alternative comes from Cypriniformes, a relatively medium sized genus known as Pangio. I’ve previously owned Pangio for many years and they are one fish I would definitely keep again.
Pangio myersi and Pangio semicincta/kuhlii
Pangio contains around 32 species (Bohlen et al., 2011), exclusive to South East Asia. They are clearly small anguilliform (the eel shape, not the taxa) but borderline very similar to the larger loach relative, Misgurnus anguillicaudatus (weather loach). For the aquarist the taxonomy can prove confusing with revisions that are not always well known such as the synonymy of Pangio semicincta and Pangio kuhlii (Kottelat, M & Lim, 1993) but is frequently debated seemingly with little explanation as to why (Eschmeyer, 2025). Molecular phylogenetics hasn’t seemed to have solved the confusion, or it’s suggested that the two species are the same (Bohlen et al., 2011). Another problem is Pangio myersi is nested within the two (Bohlen et al., 2011) although easily diagnosed for aquarists by thick barring from dorsal to abdomen (Kottelat, M & Lim, 1993).
There doesn’t seem to be immediately much morphological diversity in this genus, there is a diversity of patterning. While many will attempt diagnosing species by colouration, this has been called into question with solid marked individuals being identified as those with stripes (Bohlen et al., 2011). Like all loaches they contain small scales that to some can make them seem scaleless.
My interest is largely in morphology and like many there seems to be no anatomical studies. The majority focuses in the taxonomic records and this makes it really difficult to understand the morphology that might have ecological importance and also husbandry. We can clearly see a inferior (ventrally facing) mouth so they feed below and given the barbels it seems a common trait with those rooting in the substrate given they are not mobile.
Pangio cuneovergata
In the aquarium hobby we keep very few species but a diversity is starting to be imported and not just as bycatch. You can expect to find of the distinctively patterned species Pangio semicincta/kuhlii, P. myersi and P. shelfordii. The smallest species that is now being imported in reasonable numbers is Pangio cuneovergata. There are a few solid coloured species and these are likely Pangio oblonga and P. anguillaris, potentially also P. malayana who is shown to have solid individuals.
Pangio semincincta/kuhlii
How to Identify your Pangio?
This is a tricky topic but there are multiple sources that holds clues.
Figure 1: A Key to the Pangio of the Malay Penninsula as described in: Bohlen, J., Šlechtová, V., Tan, H. H., & Britz, R. (2011). Phylogeny of the southeast Asian freshwater fish genus Pangio (Cypriniformes; Cobitidae). Molecular Phylogenetics and Evolution, 61(3), 854-865.
As described previously there are some hints you can get from experiencing the species in captivity but there is a lot of cross referencing, where you can linking to a locality and exploration of images that could be of use. I find Bohlen et al. (2011; Fig 1) possibly the most useful of the papers for identification of species even if images are limited. One must also remember where you might have imported from one country it doesn’t mean that is where it is caught.
Habitat
There is little ecological details on these fishes. Habitat likely differs per species. Largely found in blackwater although potentially also highly turbid waters with variable or seasonal currents (Bohlen et al., 2011). Yet in the literature very little else is recorded.
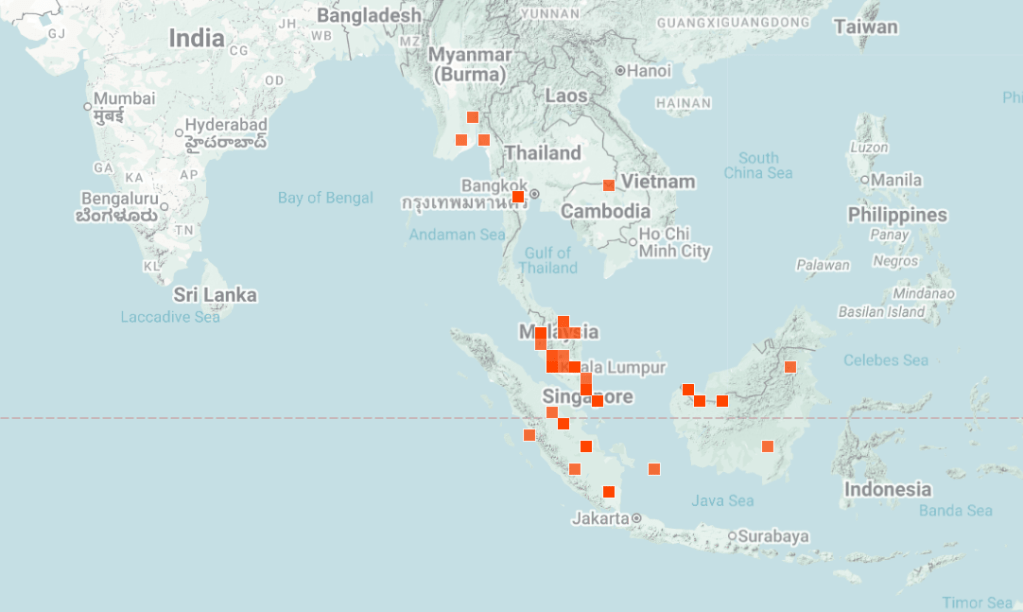
Figure 2: Geographical distribution of Pangio according to INaturalist (2025)
Some assumptions can be made from the locality of these fishes (Fig 2), particularly in reference to temperature, although you will also need to check against elevation and other factors such as is the water body sheltered and therefore cooler. INaturalist (https://www.inaturalist.org/)and GBIF (https://www.gbif.org/) can be useful here although neither make any records of environmental factors. If someone is familiar with these websites then there is other extractions of data that can be done but I am not someone who works with species distributions. Other parameters can be tricky without knowing the geology of the region, some rocks dissolve more readily whereas others allow tannins or just rainwater to drain rapidly through without dissolving minerals.
We also know Pangio almost always seem to breed in tanks with gravel, there are probably exceptions so the substrates must have gravel where they are located with the exception of maybe more elongate species who similar to lamprey bury in silt and sands.
What setup would suit Pangio?
Generally I don’t think the current ideal setup for Pangio needs any adapting, sandy areas and gravel based areas, leaf litter providing many hiding places but also wood or rocks for more solid refuges.
One of my setups in 2017 for multiple Pangio spp., filtered by a Fluval 306 (I think).
A reasonable current but it wouldn’t have to be that strong so a sponge filter would suffice. I would consider with externals, internals or similar filters whether the fishes could get in, they are notorious for finding their ways into filters so either providing an inlet guard or changing filtration method particularly for those like Pangio cuneovergata. Undergravel filters sound great and somewhat are, the fishes will make their way into them and likely also live below the grids but many do report them spawning well there.
There is no doubt these fishes come from more soft acidic water based on locality and therefore I recommend a pH of around 6-7.5 for various reasons. There is flexibility and these fishes have proven themselves very hardy in captivity. Total Dissolved Solids (TDS) below 200ppm but ideally below that 100ppm range unless your water has potentially skewed values. Temperature is definitely the tricky one without knowing exactly the temperature of their caught locality, I would personally cross reference where they are found with water temperatures of the area and who knows other species in the area might have ecological data. Generally 24-28c seems the most ideal but you could be flexible particularly on that upper end to go higher.
They are often noted as being particularly shy although I’ve found with dimmer and dappled lighting but also frequent exposure they don’t seem to be that shy and show a lot of normal feeding and explorative behaviour. Plenty of cover gives them somewhere to retreat to.
References:
Bohlen, J., Šlechtová, V., Tan, H. H., & Britz, R. (2011). Phylogeny of the southeast Asian freshwater fish genus Pangio (Cypriniformes; Cobitidae). Molecular Phylogenetics and Evolution, 61(3), 854-865.
Kottelat, M & Lim, K. K. P. (1993) A review of the eel-loaches of the genus Pangio (Teleostei:Coditidae) from the Malay Peninsula, with descriptions of six new species. Raffles of Zoology, 41(2): 203-249.
It is frequently understood that ornamental fish nutrition differs from food fish nutrition due to differing aims, yet many fish feeds use the same ingredients (Yanong, 1999; Vucko et al., 2017). Much of the research into fish nutrition no doubt due to funding focuses on aquaculture (farming fishes) the concept of efficiency, a fish farmer wants a fish to grow large in as minimal time (usually months) as possible for as little cost as possible. This opposes the fishkeepers who want a diet to give the fish longevity, colour and long term reproductive success, there is also a higher focus on the welfare element as fishes are our pets.
A large factor is there are many species of ornamental fishes across many different fish groupings, this proposes a challenge as reflected in this is the wide range of dietary niches that fishes can exhibit. There are carnivores which can be split into different invertebrate specialist, generalists and also piscivores. Herbivory which can be split into frugivores, algivores, plant eaters and many different specialist and generalist niches. Detritivory also exists and while often classified as feeding on decaying matter in the aquatic sense this could be many things from microbial matrices crossing over a lot with algivory to a mixture of unidentified matter. Omnivory does exist but it is often vague to clearly state a fish is an omnivore as there are few which broadly generalize. And while clades can be generalized such as Loricariids being largely algivores and detritivores, there are carnivores; similarly for Cichlidae the majority are carnivores but there are many exceptions verging into algivory. An important concept as above is generalism vs specialism, the degree of specialism varies between clades and this differs largely between captivity and the wild (Golcher-Benavides and Wagner, 2019). This poses a particular issue for the ornamental aquarist as many aquarium diets fall along the lines of generalism, the ingredients do not largely differ between many products and even brands. This means that even offering a range of different products doesn’t always cater for diversity or all the dietary niches of the fishes you have in the aquarium.
It is always advisable to research the wild diet of the fishes as this will give a good idea of the dietary niche a fish has evolved for over hundreds or thousands of years. Often there are clues in the morphology of the fish, such as head shape, pharyngeal jaw shape and dentition (Burress, 2016). Well known hobby ideas of fish diets can lack fact checking such as the common misconception of Symphysodon (discus fishes) being carnivores yet their wild diet follows more algivory/detritivory (Crampton, 2008) and their morphology is most similar to Tropheus (Fraser et al., 2009), a known algivore. There does remain issues with understanding of wild fish diets, there can be frequent gaps or sampling errors. Such as the original understanding behind the diets of Corydoradinae was based on aquarium fishes, not the wild fishes so it would be better to identify gut analysis and isotope studies of the wild fishes (Alexandrou et al., 2011).
What are the basic nutritional requirements of ornamental fishes?
Like any animal there are broadly the same categories here; carbohydrates, lipids, proteins, vitamins and minerals.
Proteins These are the nutritional requirements many will state of importance with fishes being high and low protein dependent. Although the idea of herbivores having a lower protein requirement then carnivores in the aquatic world fails to understand the high protein levels of algae’s opposed to the terrestrial vascular plants. Different sources of protein do have different volumes of different amino acid and this is important to recognize. This is arguably more important then the actual volume of protein along with accessibility of the protein source. Algivorous fishes might have a lower requirement for protein (Vucko et al., 2017; Yanong, 1999). A large factor of protein requirements is the fishes age, temperature and genetics, as with any nutrient all juvenile fishes will require higher nutrition then those whose growth rate has plateaued more. Excessive protein also increases ammonia production.
Carbohydrates are used as an energy source for many fishes. It is understood that starch unlike cellulose is the utilized form for many ornamental fish feeds, many herbivores and omnivores do not require carbohydrates. Excessive carbohydrates can prevent protein and sugar uptake (Yanong, 1999).
Lipids are another energy source and important for ornamental fishes (Vucko et al., 2017; Yanong, 1999) particularly for spawning fishes who will require them for egg development.
Vitamins and minerals are a complex and diverse topic given how many are vital for life within fishes, without them it can lead to many health issues. I would encourage the reader to read Yanong (1999; https://www.researchgate.net/publication/12102293_Nutrition_of_Ornamental_Fish) for details into this. Some of these vitamins and minerals are taken through diet and others through water, when it comes to water uptake this must be considered with the wild water parameters of the fishes as where calcium is low in the water uptake must be largely from diet. The mineral content of diets can be vague although the total volume is reflected in what is called ash. No, ash isn’t literal ash it means mineral content as when you burn down a diet all that is left is the minerals.
Colour enhancers, this is a concept that is frequently mentioned in food reviewed largely in reference to a diversity of carotenes. While it will always be stated in marketing and social media as a benefit to a fish feed, most fish feeds contain red enhancers of some kind, this is largely a benefit to most fishkeepers excluding those with yellow discus who will change their colour when feeding on it.
Largely these requirements are so complex it’s tricky to address them with one diet when we keep so many species.
Why might change be needed in what we feed our fishes?
Seeing as many farmed fishes diets do not replicate their wild diets, many do not replicate a diversity in fish diets that is the start. Maybe sometimes they do in the labels but the ingredients do, when a diet aimed at algivores has the same ingredients as one aimed at carnivores. For algivores we already know just one algae can compete with a commercial mix of many ingredients which says a lot (Vucko et al., 2017).
It has become clear that diet is key in the health of a fish with diet influencing the development of tumours (cancer; Spitsbergen et al., 2012), liver damage (Rašković et al., 2011) and other health conditions (Žák et al., 2022). There is the potential of poor fecundity and reproductive lifespans on certain diets although could require further exploration. Many fishes we just don’t see them grow to their adult sizes let alone spawn and mortality might be high where they can refuse to feed on traditional captive diets.
References:
Alexandrou, M. A., Oliveira, C., Maillard, M., McGill, R. A., Newton, J., Creer, S., & Taylor, M. I. (2011). Competition and phylogeny determine community structure in Müllerian co-mimics. Nature, 469(7328), 84-88.
Burress, E. D. (2016). Ecological diversification associated with the pharyngeal jaw diversity of Neotropical cichlid fishes. Journal of Animal Ecology, 85(1), 302-313.
Crampton, W. G. (2008). Ecology and life history of an Amazon floodplain cichlid: the discus fish Symphysodon (Perciformes: Cichlidae). Neotropical Ichthyology, 6, 599-612.
Fraser, G. J., Hulsey, C. D., Bloomquist, R. F., Uyesugi, K., Manley, N. R., & Streelman, J. T. (2009). An ancient gene network is co-opted for teeth on old and new jaws. PLoS biology, 7(2), e1000031.
Golcher-Benavides, J., & Wagner, C. E. (2019). Playing out Liem’s paradox: opportunistic piscivory across Lake Tanganyikan cichlids. The American Naturalist, 194(2), 260-267.
Vucko, M. J., Cole, A. J., Moorhead, J. A., Pit, J., & de Nys, R. (2017). The freshwater macroalga Oedogonium intermedium can meet the nutritional requirements of the herbivorous fish Ancistrus cirrhosus. Algal research, 27, 21-31.
Rašković, B., Stanković, M., Marković, Z., & Poleksić, V. (2011). Histological methods in the assessment of different feed effects on liver and intestine of fish. Journal of Agricultural Sciences (Belgrade), 56(1), 87-100.
Spitsbergen, J. M., Buhler, D. R., & Peterson, T. S. (2012). Neoplasia and Neoplasm-Associated Lesions in Laboratory Colonies of Zebrafish Emphasizing Key Infl uences of Diet and Aquaculture System Design. Ilar Journal, 53(2), 114-125.
Yanong, R. P. (1999). Nutrition of ornamental fish. Veterinary Clinics of North America: Exotic Animal Practice, 2(1), 19-42.
Žák, J., Roy, K., Dyková, I., Mráz, J., & Reichard, M. (2022). Starter feed for carnivorous species as a practical replacement of bloodworms for a vertebrate model organism in ageing, the turquoise killifish Nothobranchius furzeri. Journal of Fish Biology, 100(4), 894-908.
I commonly get asked what I’d feed different groups of pleco’s and it does vary by the pleco, Loricariid but also by what is available in your country. Some genera are more generalist and forgiving then others, some will withstand a less then ideal diet for maybe a few decades and others a few months/days.
While I would love to provide more details some I cannot list yet or am not entirely clear on some aspects of a species dietary ecology. So this should hopefully help for now.
I shall split it up by diet, some species might feed on a different diet to what you expect so please search for the genus/species.
The majority of Loricariids are algivores and detritivores so this contains the larger amount of categories.
Algivores
This is one of the largest categories as many Loricariids specialize in different algae’s, biofilms etc. But due to mode of feeding and availability of ingredients I will kind of have to generalize. Due to this the algae growing in the aquarium might be the wrong species so the fish might not feed on it, regardless any algae in the aquarium will not sustain most species long term.
Prepared diets:
Repashy Super Green
Repashy Soilent Green (you can add in extra algae powders to bulk it out, fishes seem to prefer this diet).
Making your own Algivore Diet
This is a trial as it seems Repashy is becoming unavailable in many countries. I have done many different trials and tests but am still developing something.
Nutritional ingredients:
These are the main ingredients and should make up at least 80% of the nutritional ingredients used but can makeup the whole diet minus gelling agents (Vucko et al., 2017). Percentages might vary, ideally try to include higher volumes of those highlighted in bold, not all will be available so try to include as wide of a diversity of possible. High spirulina content might take a while for the fishes to get used to. Seaweeds will need to be blended or might not be eaten.
Chlorella algae (Vital)
Spirulina algae (Vital)
Seaweed meal (Vital)
Kelp meal (vital)
Wakame algae (vital)
Nori (Vital)
Bladderwrack
Other human consumption seaweeds and algaes.
Potentially mosses, never tried but are recorded in Loricariid diets.
Herbal ingredients:
These should be very limited excluding the mushrooms I wouldn’t go above around 1-2% per ingredient.
Paprika, associated with red enhancing.
Mushrooms, dried or powdered while not entirely known it is potential they feed on fungi in the wild. While edible mushrooms might not be the same taxa it does seem to be a taste enhancer for fishes at least.
Basil, associated with improved physiological and immunological health while being an attractant (Mansour et al., 2023).
Ginger, feed attractant and immunological support (Ahmad et al., 2024).
Garlic, I don’t always use it but feed attractant with potential immunological support but can cause liver damage.
Seeds, particularly found in the guts of Hypancistrus and potentially Peckoltia. Higher in fats and proteins.
Gelling agents:
I would recommend using carrageenan powder due to it lasting longer then the alternatives. I would use this regardless as to whether it is a carnivorous or herbivorous diet.
Who are the algivores that we keep?
Ancistrus
Baryancistrus
Dekeyseria
Farlowella
Hemiancistrus
Hypostomus
Isorineloricaria
Lamontichthys
Lasiancistrus
Nannoptopoma
Otocinclus
Panaque
Panaqolus
Parancistrus
Parotocinclus
Pseudancistrus
Pseudorinelepis
Pterygoplichthys
Rhinotocinclus
Spectracanthicus zuanoni/punctasissimus
Sturisoma/Sturisomatichthys
This diet will cover most of Loricariidae but particularly these genera, while they might be also more detritivorous in the wild this is the closest we can get to their natural diet.
Regarding Hypancistrus, Peckoltia and potentially Panaqolus I would add seeds to their diet and maybe look at the addition of infrequent invertebrates.
A little more carnivorous
If you want to increase the volume of carnivorous ingredients, Loricariids don’t consume fishes in the wild so we will be looking at invertebrates. Due to the presence of thiaminase in some ingredients I do not recommend the frequent use of mussels or prawns.
So as you’re looking more into carnivory I would increase the volume of these ingredients, luckily for carnivorous ingredients you could feed as a frozen or live food they have to forage.
Ingredients:
Daphina
Brine shrimp
Tubifex
Bloodworm (Chironomatid larvae).
Cockels
Mysis
Red plankton
Ant eggs
Earthworms
Cyclops
Whiteworms
Blackworms
Vinegar worms
What about molluscivores?
Scobiancistrus, Leporacanthicus, Pseudohemiodon, Loricaria and Planiloricaria are likely capable on feeding on mussels and occasionally this is proven. A diversity of snails can be trialed for the Scobinancistrus and Leporacanthicus larger species such as Ampulluridae would be ideal as these are evolved to extract snails from their shells, escargot snails that are not treated with garlic would be interesting to explore. For others then smaller snails whether it be juveniles of harder species of pest snails.
Plant eaters
Realistically many Loricariids do not consume traditional plants so often these are best to identify if a fish is feeding or not. Some very broad generalist taxa might consume more like Pterygoplichthys and Hypostomus.
Using other premade foods as a base.
This is largely only possible with gel diets but possibly some pastes. The main rule is not to add so many that the gelling agent doesn’t hold as well as it used to but also this will depend on how fast your fishes feed.
What base diets can you use?
Repashy. It does have a wide range of other ingredients.
In the Bag Tropical Fish UK’s pleco pops. Very strong gelling agents and true carnivorous and herbivorous diets.
EBO pastes
Tropical’s gels/pastes
Testing diets to identify if they are being used.
Glass petri dishes can be ideal here as they sink and are inert, you can pipette or place food on and ideally it will not be disturbed over the time you are not observing the food being eaten. Therefore it can be a reliable method of identifying what is taken and what is not.
References:
Ahmad, I., Irm, M., Ahmed, I., Haoran, Y., Taj, S., Bhat, T. A., … & Amin, A. (2024). Role of ginger in fish nutrition with special emphasis on growth, health, gut and liver morphology. Journal of the World Aquaculture Society, 55(6), e13101.
Mansour, A. T., Diab, A. M., Khalil, R. H., Eldessouki, E. A., El-Sabbagh, N., Elsamannoudy, S. I., & Younis, N. A. (2023). Physiological and immunological responses of Nile tilapia fed dietary supplementation of sweet basil ethanolic and aqueous extracts. Frontiers in Marine Science, 9, 1064455.
Vucko, M. J., Cole, A. J., Moorhead, J. A., Pit, J., & de Nys, R. (2017). The freshwater macroalga Oedogonium intermedium can meet the nutritional requirements of the herbivorous fish Ancistrus cirrhosus. Algal research, 27, 21-31.
Hypancistrus have long been an issue for hobbyists and taxonomists providing challenges to identify and define what is a species, over time a few have been described but leaving one of the most common species.
This topic is tricky for scientists regarding how a species is defined and where do you draw a line, even trickier for hobbyists. The Rio Xingu species have been particularly tricky as there are many striped species with only Hypancistrus zebra being particularly distinctive. For the hobbyist the L number system can add to the confusion as while different individuals can be given different L number it doesn’t infer they are different species. Morphology can be tricky to navigate as there are many very diverse species both morphologically and genetically for example Baryancistrus xanthellus (of which does include a green variant, verde that is not B. chrysolomus) or Peckoltia sabaji (Fig 1; Magalhães et al., 2021; Armbruster 2003).
Figure 1: The morphological diversity of Baryancistrus xanthellus, the gold nugget pleco (L177, L018, L081, L085, verde) as featured in Magalhães, K. X., da Silva, R. D. F., Sawakuchi, A. O., Gonçalves, A. P., Gomes, G. F. E., Muriel-Cunha, J., … & de Sousa, L. M. (2021). Phylogeography of Baryancistrus xanthellus (Siluriformes: Loricariidae), a rheophilic catfish endemic to the Xingu River basin in eastern Amazonia. Plos one, 16(8), e0256677.
Some of this morphological and genetic diversity can be based on different populations and localities, it is tricky to infer whether there is interbreeding as to when and extent this occurs without detailed analysis for both morphologically and genetically. We also risk drawing lines between populations or individuals of the same species that don’t exist in nature.
Commonly understood is the importance of species as a biological unit and in some manner it is, but this is no one overarching definition for a species, it’s much more complex then that. As said earlier species can be both morphologically and genetically diverse or not at all, it varies so much and on where the line is drawn. The common misconception is that genetics solves any issues with defining a species but when you create these trees to plot species different genes, regions or even whether you use mitochondrial or nuclear DNA can infer different groupings. But this reliance on species being the important factor that matters for many aquarists ignores much of this and can lead to splitting species into unrealistic groupings. Realistically like the killifish and Poecilidae sides of the hobby, we need to recognize populations are as valuable as species, even if they cross or not. Populations might have unique genetics or morphology, doesn’t make them different species but we should really think through how we breed our fishes and what individuals we choose. If fishes come from different suppliers maybe double checking locality, maybe considering if certain captive bred fishes are useful for maintaining a population.
So in summary just because some species might look different it doesn’t mean they are but doesn’t mean they aren’t distinct populations that shouldn’t be valued.
Description for Hypancistrus seideli and H. yudja:
Sousa LM, Sousa EB, Oliveira RR, Sabaj MH, Zuanon J, Rapp Py-Daniel L. (2025). Two new species of Hypancistrus (Siluriformes: Loricariidae) from the rio Xingu, Amazon, Brazil. Neotropical Ichthyology. 23(1). https://doi.org/10.1590/1982-0224-2024-0080
These exciting descriptions help us understand the Loricariids we keep in the aquariums better and more accurately describe them. Hopefully it leads to further studies of Hypancistrus.
Figure 2: Hypancistrus seideli as featured in: Sousa LM, Sousa EB, Oliveira RR, Sabaj MH, Zuanon J, Rapp Py-Daniel L. (2025). Two new species of Hypancistrus (Siluriformes: Loricariidae) from the rio Xingu, Amazon, Brazil. Neotropical Ichthyology. 23(1). https://doi.org/10.1590/1982-0224-2024-0080
This species includes the L numbers: L333, L066, L236, L287, L399, L400.
This species includes the common names: King tiger pleco, maze zebra pleco.
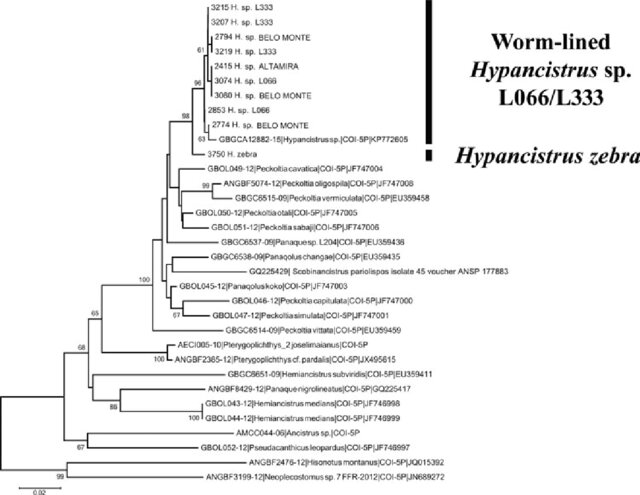
Diagnosed by alternating dark and pale vermiculation’s from currently described species although recognised as extremely varied (Sousa et al., 2025). Hypancistrus seideli covers a wide range of the Hypancistrus diversity in the Rio Xingu and some of the most popular species in the aquarium trade. Although morphologically diverse (Fig 2) there it seems to not have the same amount of molecular diversity so further inferring at least L066 and L333 regardless are the same species. Phylogenetically there also seems to be an issue to designate them as different species given L066 and Belo Monte seem to be paraphyletic (Cardoso et al., 2016). Although using sequences from a public database does rely on correct identification of those sequencing the samples (Fig 3).
Figure 3: Molecular phylogeny using the COI barcode sequences located from a public database as featured in: Cardoso, A. L., Carvalho, H. L. S., Benathar, T. C. M., Serrao, S. M. G., Nagamachi, C. Y., Pieczarka, J. C., … & Noronha, R. C. R. (2016). Integrated cytogenetic and mitochondrial DNA analyses indicate that two different phenotypes of Hypancistrus (L066 and L333) belong to the same species. Zebrafish, 13(3), 209-216.
Etymology: Hypancistrus seideli is named after the well known and respected aquarist Ingo Seidel who has contributed a lot to the knowledge of Hypancistrus (Sousa et al., 2025).
Habitat: While the paper doesn’t go into detail that isn’t well known it describes their environment as rocky with strong currents (Sousa et al., 2025).
Figure 4: Hypancistrus yudja as described in: Sousa LM, Sousa EB, Oliveira RR, Sabaj MH, Zuanon J, Rapp Py-Daniel L. (2025). Two new species of Hypancistrus (Siluriformes: Loricariidae) from the rio Xingu, Amazon, Brazil. Neotropical Ichthyology. 23(1). https://doi.org/10.1590/1982-0224-2024-0080
This species includes the L numbers: L174.
This species includes the common names: Ozelot pleco.
Diagnosed by large brown splotches and saddles on a tanned background (Sousa et al., 2025).
Etymology: Named after the Yudjá people of the Volta Grande, Rio Xingu, Brazil who are located in the same area as these fishes and described as equally threatened by the Belo Monte dam (Sousa et al., 2025).
Habitat: Located specifically from deep but rocky waters but remains hidden in crevices for large amounts of time (Sousa et al., 2025).
Armbruster, J. W. (2003). Peckoltia sabaji, a new species from the Guyana Shield (Siluriformes: Loricariidae). Zootaxa, 344(1), 1-12.
Cardoso, A. L., Carvalho, H. L. S., Benathar, T. C. M., Serrao, S. M. G., Nagamachi, C. Y., Pieczarka, J. C., … & Noronha, R. C. R. (2016). Integrated cytogenetic and mitochondrial DNA analyses indicate that two different phenotypes of Hypancistrus (L066 and L333) belong to the same species. Zebrafish, 13(3), 209-216.
Magalhães, K. X., da Silva, R. D. F., Sawakuchi, A. O., Gonçalves, A. P., Gomes, G. F. E., Muriel-Cunha, J., … & de Sousa, L. M. (2021). Phylogeography of Baryancistrus xanthellus (Siluriformes: Loricariidae), a rheophilic catfish endemic to the Xingu River basin in eastern Amazonia. Plos one, 16(8), e0256677.
Identifying Corydoradinae Catfish by Ian Fuller and Hans Evers has been long awaited by many catfish enthusiasts whether Corydoradinae is their interest or other Siluriformes. Using the vital recent revision of the genus (Dias et al., 2024) we now have an accompanying book accessible to hobbyists but also enthusiasts.
The revision of Corydoradinae was long needed as in some way resurrecting previous genera and reflecting the previous lineage system hobbyists used, this lineage system was also reflected in the fishes phylogeny/evolution (Dias et al., 2024). Revising the genus Corydoras and subfamily Corydoradinae reflects the morphology/anatomy and ecology better and helps us understand how to keep our Corydoradinae better.
This book offers a modern view at the group brimming with lifetimes of experience and knowledge. I need not introduce the authors as both are extremely well respected not just within Corydoradinae, catfishes but the hobby in general. This collaboration creates the perfect all round collaboration.
The science is always useful to really understand aquariums and how we keep our fishes of which some sources very much lack. Identifying Corydoradinae includes some fascinating science you wont see included elsewhere that is described in an easily understood manner to someone who might not be trained in the sciences. One of the most interesting topics was genome expansion written by Professor Martin Taylor, a scientist who studies the genetic aspects of Corydoradinae. I also very much enjoyed the section on toxins or poisons within Corydoradinae from Professor Eric Thomas as this is at the cutting edge of science, it’s a topic that is not well understood yet this book offers the latest knowledge as to the topic. For me as having an interest in ecology I was really keen to see mentions as to the fishes ecology and habitats. The ecology section was very useful for hobbyists in describing actually where these fishes are found from experience and knowledge, giving the hobbyist an idea of how to keep the fishes and breed them. Diet has sometimes been misunderstood in the hobby and the inclusion of what we know these fishes feed on is no doubt a benefit. Dr. Luiz Tencatt goes into detail on these fishes diets and hopefully this helps hobbyists know what to feed their fishes.
The layout of the book is useful for the hobbyist as logically passes through the genera before the described species and then the undescribed. This means the hobbyist can quickly identify the species they are most interested in at the time. Brochis is additionally split up which helps identify the various lineages, not completely removing the lineage system therefore the usefulness of it but that would require another scientific revision.
All the species are clearly explained with distributions, size and also detail as to their identification and ecology. To help the enthusiast who might not be too clear on the various genera there is a guide to the various genera and the key features a hobbyist can use to identify them. Plenty of clear images are provided of not just the adults but variants and as the fish ages.
In conclusion, this is a great book with so much detail for any catfish enthusiast. It provides information to help the hobbyists understand a large group of catfishes that can be tricky to identify. Certainly a book you can read in one session but for many years will be of great use.
References:
Dias, A.C., Tencatt, L.F., Roxo, F.F., Silva, G.D.S.D.C., Santos, S.A., Britto, M.R., Taylor, M.I. and Oliveira, C., 2024. Phylogenomic analyses in the complex Neotropical subfamily Corydoradinae (Siluriformes: Callichthyidae) with a new classification based on morphological and molecular data. Zoological Journal of the Linnean Society, p.zlae053.
While it is not always obvious Loricariids all have teeth and they show a wide range of morphological disparity (Lujan & Armbruster, 2012). Morphological disparity refers to that range of different anatomy within a group. Unlike many fishes Loricariids are rarely gape limited, their prey (that includes algae) is not limited by the size of their mouth and this makes comparison with traditional fishes like carp or cichlids limited.
Figure 1: Pterygoplichthys joselimaianus jaw, a typical jaw for Loricariidae.
Loricariids feed largely by a rasping motion, this is extremely similar to how snails feed. Snails also have teeth on a ribbon like organ known as a radula (Fig 2).
Figure 2: Snail body plan including the mouth anatomy. Unknown source.
Both snails and Loricariids use their jaws, containing the teeth to basically scrape at a surface (rasp), it can be the food item itself but it could be rocks or wood to extract food.
Figure 3: Leporacanthicus joselimai
While I say the majority of Loricariid jaws are similar to snails not all are, there are carnivorous genera such as Pseudohemiodon or Scobinancistrus who differ in how they move and function. Carnivorous genera have elongate fewer teeth with often narrower oral jaws but can be much more robust (Fig 3), or in some those oral jaws are almost entirely just the jaws, reduced tooth cups. It’s easy to say these carnivores are using different morphology for the same solution to carnivory but maybe in a different place, one feeds amongst crevices (those with the elongate teeth and jaws) and others amongst the substrate (those with reduced jaws). We do have a slight exception with Spatuloricaria, an obvious substrate feeder but it seems to use the substrate a little differently and feed on different invertebrates.
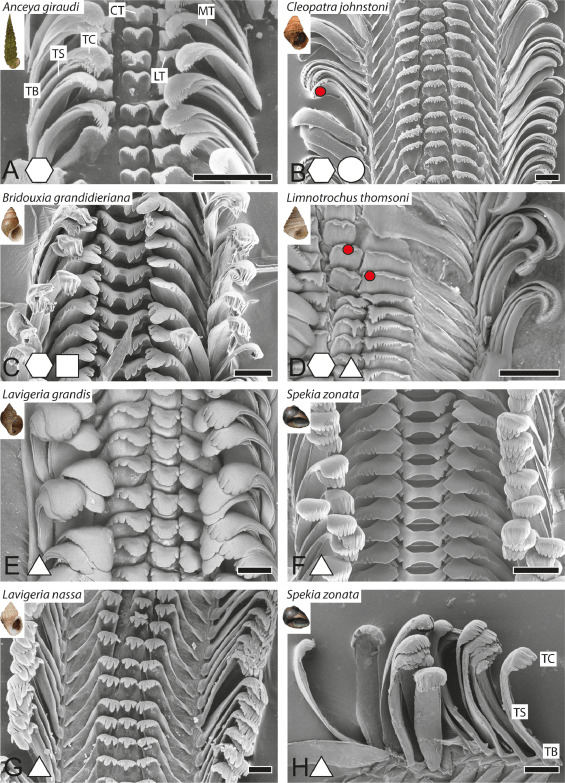
Figure 4: Gastropod radula diversity. Krings, W., Kovalev, A., & Gorb, S. N. (2021). Collective effect of damage prevention in taenioglossan radular teeth is related to the ecological niche in Paludomidae (Gastropoda: Cerithioidea). Acta Biomaterialia, 135, 458-472.
Snails, Gastropods have long been studied in terms of their radula diversity (Fig 4), I assume this is due to the fact when you’re dealing with preserved snails there are fewer tissues to identify the species. Additionally they make great models for understanding how anatomy relates to morphology, ecomorphology. Gastropods are everywhere and it’s easy to find those that scrape algae’s off rocks vs more carnivorous gastropods. To put it simply though, Gastropods feed by rasping and their teeth are uniquely shaped to what they are feeding on.
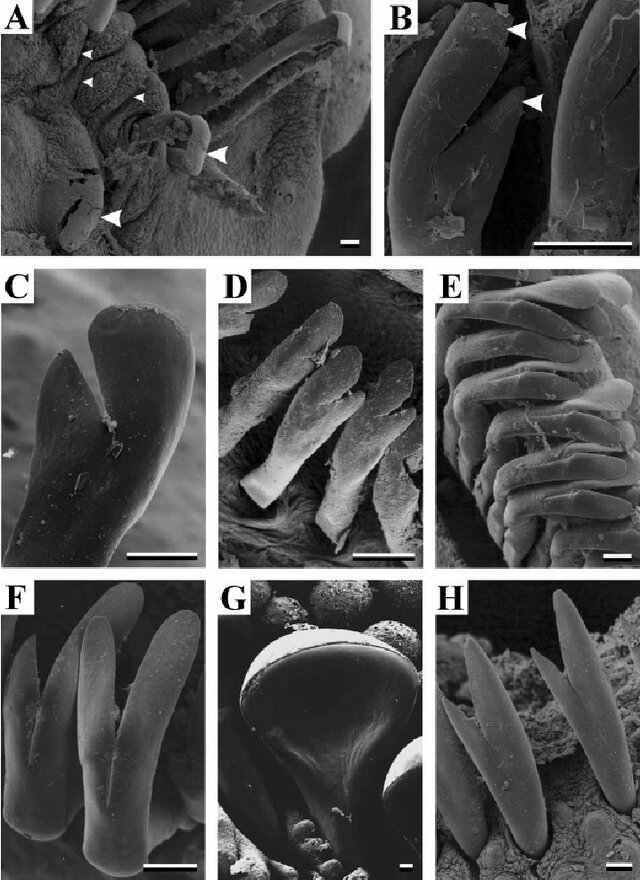
I think even just ignoring carnivory Loricariidae shows a wide diversity of tooth morphological disparity but there is little studies regarding that in relation to their ecology. Plenty of these studies focus on the development and morphology (Geerinckx et al., 2007). What there is is a fascinating study looking at another part of the fishes anatomy that could be similar, the unculi, small protrusions on the oral disc’s of the fishes. While the study focuses on how these structures allow for the fishes to inhabit certain habitats, could these also function in a similar fashion to radula?
Figure 5: The diversity of Loricariid teeth, Geerinckx, T., De Poorter, J., & Adriaens, D. (2007). Morphology and development of teeth and epidermal brushes in loricariid catfishes. Journal of morphology, 268(9), 805-814.
The diversity of Loricariid tooth morphological diversity is clear (Geerinckx et al., 2007) and we clearly see that Loricariids have a diversity of diets beyond herbivory and carnivory (Lujan et al., 2012), whatever they really mean to aquatic animals.
When looking outside of carnivory there is clear differences in morphology, none are so much clearer then those Loricariids that utilize wood. These genera display clearly spoon shaped teeth even if these genera (Panaqolus, Panaque, Hypostomus cochliodon group etc.) do not digest the wood and it is simply where they might find food. Compared with carnivores such as Leporacanthicus, these have more elongate teeth but it depends on where they are accessing their food. This difference is also reflected in gastropods whether they be snails or slugs have evolved teeth on their radula that reflect not just their diet but the methods they use to extract it. Elongate pointed teeth infers carnivory whereas further cusps leans towards herbivory. Perhaps carnivory requires less complexity to herbivory and I assume largely as carnivory relies on more then the teeth to extract food.
References:
Geerinckx, T., De Poorter, J., & Adriaens, D. (2007). Morphology and development of teeth and epidermal brushes in loricariid catfishes. Journal of morphology, 268(9), 805-814.
Krings, W., Konn-Vetterlein, D., Hausdorf, B., & Gorb, S. N. (2023). Holding in the stream: convergent evolution of suckermouth structures in Loricariidae (Siluriformes). Frontiers in Zoology, 20(1), 37.
Krings, W., Kovalev, A., & Gorb, S. N. (2021). Collective effect of damage prevention in taenioglossan radular teeth is related to the ecological niche in Paludomidae (Gastropoda: Cerithioidea). Acta Biomaterialia, 135, 458-472.
Lujan, N. K., & Armbruster, J. W. (2012). Morphological and functional diversity of the mandible in suckermouth armored catfishes (Siluriformes: Loricariidae). Journal of Morphology, 273(1), 24-39.